

67 гостей
VIVOS VOCO: Е.Б.Абрамова, В.Л.Карпов, "Протеасома: РУЙНУВАННЯ В ІМ'Я ТВОРЕННЯ"
© Е.Б.Абрамова, В.Л.Карпов
протеасома:
РУЙНУВАННЯ В ІМ'Я ТВОРЕННЯ
Є.Б. Абрамова, В.Л. Карпов
Олена Борисівна Абрамова, к. Б. н., ст. наук. співр. Інституту біології розвитку ім. Н.К.Кольцова РАН.
Вадим Львович Карпов, д. Б. н., проф., зав. лаб. Інституту молекулярної біології ім. В.А.Енгельгардта РАН.
Відомо, що в живій клітині містяться безліч різних білків. Одні з них існують досить довго, інші живуть від декількох хвилин до 2-3 ч. Останні синтезуються тільки в певний момент життя клітини, у відповідь на деякі внутрішні і зовнішні імпульси. Коли потреба в такому білку відпадає, спеціальні фактори сигналізують про те, що його синтез повинен бути зупинений. Діють вони або на етапі зчитування матричних РНК (мРНК) з ДНК (фактори транскрипції), або на стадії синтезу білка з уже наявних РНК-матриць. Однак, якщо якийсь білок існує, але в його функціонуванні клітина більше не потребує, повинен бути і механізм, що забезпечує зупинку його роботи. Такий механізм давно і вельми детально досліджений для білків-ферментів. Щоб "вивести з ладу" фермент, його активність можна зупинити пригнічується (відзначено зниження) речовинами білкової або небілкової природи. Однак клітка впродовж свого життя синтезує і безліч білків, що не володіють ферментативною активністю, але тим не менш "виконуючих" різноманітні ролі. Такі білки не тільки припиняють свою роботу в певний момент, але й існують дуже недовго (тому й називаються короткоживущими) в порівнянні з часом життя клітини.
Очевидно, що якимось чином повинні віддалятися і "зробили свою справу" білки, інакше вони переповнять клітку і зруйнують її. Може бути, вона справляється з ними за допомогою добре вивченого протеолізу (розпаду білків під дією спеціальних ферментів)? Але тоді незрозуміло, чому не ушкоджуються при цьому структурні компоненти клітини і ще потрібні їй білки. Протеолиз в лізосомах - процес неспецифічний, в цих освічених мембраною "мішечках" з набором ферментів гідролаз молекули білків розщеплюються до амінокислот, що йдуть потім в обмінні процеси клітини (тут же гідролізуються нуклеїнові кислоти і полісахариди). У вищих евкаріот лізосоми руйнують тільки білки, пов'язані з мембранами, а також чужорідні, захоплені під час ендоцитозу (наприклад, вірусні або бактеріальні).
До початку 80-х стало ясно, що ефективна регуляція не тільки кількості, але і функції багатьох білків залежить і від процесів, пов'язаних з деградацією.
багатоликі частки
В цей же час в клітці був виявлений високомолекулярний білковий комплекс, який працював в певних умовах як кілька протеолітичних ферментів [ 1 ]. Однак, що дуже важливо, свою руйнівну активність комплекс виявляв тільки в явно нефизиологических умовах під дією змінюють його структуру речовин. Він був знайдений в клітинах як найпримітивніших, так і вищих евкаріот, причому і в ядрі, і в цитоплазмі. Це свідчило про абсолютну необхідність комплексу для нормальної життєдіяльності клітини.
Його почали інтенсивно вивчати в багатьох лабораторіях світу, і навіть виникло безліч назв, з яких зараз найчастіше використовується той, що дали К.Танаку і А.Голдберг, - протеасома, тобто частка (сома) з протеолітичної функцією [ 2 ]. Треба сказати, що Протеасома названі дві частинки різної складності будови. Вони відрізняються також молекулярної масою і коефіцієнтом седиментації (його висловлюють в одиницях Сведберга і позначають буквою S при числі).
У 90-х роках з'ясувалося, що спочатку виділений комплекс з молекулярною масою близько 700 кДа і коефіцієнтом седиментації 20S як протеолітичного ядра входить до складу ще більш складною частки. Першу стали називати 20S Протеасома, другу - 26S Протеасома.
Потім обидві частки були виділені в очищеному вигляді, проведено рентгеноструктурний аналіз 20S протеасоми, а методами електронної мікроскопії та комп'ютерної томографії отримані "портрети" і цієї, і іншої частинки. Судячи з рентгеноструктурньїм даними, 20S протеасома являє собою порожнистий циліндр довжиною 15-17 нм і діаметром 11-12 нм, утворений чотирма лежать один на одному кільцями. Кожне з них складається з семи білкових субодиниць (молекулярною масою 20-35 кДа), причому периферичні кільця сформовані субодиницями a-типу, а два центральних - b-типу.
Канал всередині циліндра, розширюючись, утворює три камери: велику центральну і дві менші, по краях. У центральній камері і здійснюється протеоліз. Роль a - і b -колец в роботі протеасоми різна. Так, субодиниці a -кільця за рахунок своїх гідрофобних ділянок закривають отвір в центральний канал і перешкоджають випадковому проникненню білків в протеолитическую камеру. Крім того, ці ж субодиниці відповідають за приєднання інших високомолекулярних комплексів, які регулюють роботу 20S протеасоми.


Комп'ютерна томографія двох форм протеасоми [ 8 ].
Як виникає настільки складна система? Спочатку автокаталитически утворюється семічленную кільце з a-субодиниці. Потім у взаємодію з ним вступають три b-субодиниці, одна з яких несе так званий лідерних пептид, що працює як шаперон (цей поліпептид-помічник "стежить" за правильністю укладання білкової молекули). Три перших b-субодиниці визначають посадку інших, і в результаті утворюється 13S комплекс з двох різнойменних кілець. Після димеризации 13S комплексу виникає 16S попередник з чотирьох кілець, а "зріла" 20S протеасома - після автокаталитического відщеплення лідерних пептиду і вивільнення N-кінцевого треоніну, що грає найважливішу роль у формуванні каталітичних центрів.
Ми згадували, що центральна камера - це місце, де відбувається протеоліз. Саме там, на внутрішній поверхні, розташовуються каталітичні центри - три основних і два додаткових. Один з основних каталізує протеоліз за типом химотрипсина, інший - подібно трипсину, третій працює за принципом каспаз. Ці основні області локалізовані на трьох різних субодиниць кожного b -кільця, там же знаходяться і додаткові. Структура активного центру b-субодиниці незвичайна: як згадувалося, в якості основного структурного елементу, який проводить каталіз, в ньому присутня N-кінцевий треонин.
Отвір, що веде в центральний канал і утворене a-субодиниці 20S протеасоми, за розміром можна порівняти з a-спіраль білка, а крім того воно закрите їх гідрофобними N-кінцевими ділянками. Це і не дозволяє цитоплазматическим білкам проникати до місця можливого руйнування. Тому 20S протеасома, виділена в умовах, що зберігають її цілісність, гідролізує тільки короткі поліпептиди в реакції, яка не вимагає АТФ як джерела енергії. Але ж ми говорили, що головна турбота 20S протеасоми - розщеплення НЕ пептидів, а білків. Як же вони проникають в каталітичні області, якщо вхід закритий?
Все з'ясувалося: якщо використовувати агенти, здатні впливати на структуру 20S протеасоми, протеолітична активність збільшується, так як отвір в a -Кільця, провідне в центральний канал, розширюється (про це свідчать останні рентгеноструктурні дані). В експерименті таке розширення відбувається в нефизиологических умовах, під впливом низькомолекулярних речовин, а в клітці роль "откривалок" виконують макромолекулярні регулятори (активатори) досить складної будови. Поки їх відомо тільки два - РА700 і РА28 (абревіатура англ. Protein Activator). На протеолітичну активність 20S протеасоми вони впливають по-різному.
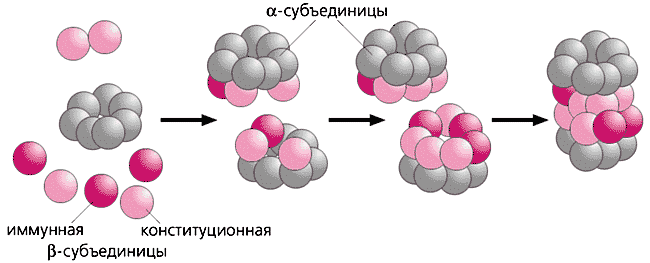
Збірка 20S протеасоми.
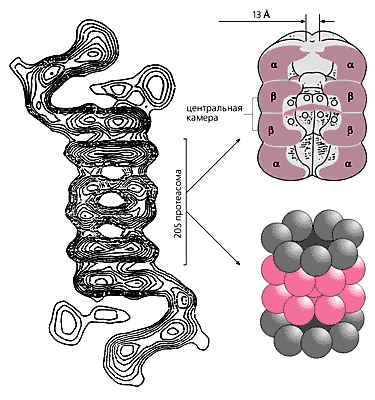
Схематична будова 26S протеасоми і протеолітичних камер.
Завдяки першому регулятору (РА700) вона бере участь в деградації основної маси клітинних білків. Формують його 17-18 субодиниць гетерогенні за молекулярною масою, амінокислотноїпослідовності і третинної структурі. У складі цього комплексу можна виділити два структурні елементи, теж аж ніяк непростих. В англомовній літературі їх називають base- і lid-complexes, що по-російськи можна передати як нижній і верхній комплекси. Так ось, нижній (базовий, опорний) приєднується до a -Кільця протеолітичного ядра і відкриває білковим субстратів доступ в каталітичну камеру. Цей елемент регулятора складається з дев'яти субодиниць, причому шість - це ферменти, що гідролізують АТФ, тобто вони поставляють енергію, необхідну на ряді етапів деградації білків. Верхній елемент побудований з восьми субодиниць, що відповідають за розпізнавання і підготовку білкових субстратів до розщеплення, за взаємодію з деякими іншими клітинними білками.
Регулятор РА700 приєднується до обох кінців протеолітичного ядра-циліндра (ця реакція також потребує енергії, її джерелом служить АТФ) і утворює дзеркально симетричну структуру, схожу на гантелі.
Каталітичне ядро (тобто 20S протеасома), пов'язане з двома регуляторами РА700, і є 26S протеасома, її молекулярна маса становить понад 2.5 МДА. Цей комплекс позначають і як РА700-20S-РА700.
Інший регулятор, РА28, теж складається з декількох субодиниць, подібно РА700, здатний приєднуватися до кінців протеолітичного ядра-циліндра з утворенням частинок РА28-20S-РА28. За рахунок цього канал в a -Кільця ядра відкривається, але гідроліз білків не йде, розщеплюються тільки короткі поліпептиди.
Зауважимо, що з 20S Протеасома можуть зв'язатися і обидва регулятора відразу, тоді виникає гібридна форма РА700-20S-РА28. Як будуть розподілені різні форми і в якій кількості знадобляться клітці, залежить від її молекулярних потреб в певний час її життя і в конкретних обставинах. Про це ми ще розповімо, а зараз повернемося до 26S Протеасома, вірніше, до її субстратів - білків.
необхідний ярлик
Отже, молекулярна машина зібрана і готова працювати - руйнувати відпрацювали білки, щоб клітина не тільки не заповнилася ними вщерть, а й використовувала білкові оскільки для своїх цілей.
У ссавців до 90% клітинних білків (не тільки всіх короткоживучих, але і більшості довгоживучих) піддається гідролізу в порожнині протеасоми. Однак, перш ніж почнеться цей процес, вона повинна розпізнати об'єкт протеолізу по якомусь ознакою, ярлику. Виявилося, маркуванням займається спеціальна система
Різні форми протеасоми і її активаторів.
ферментів (її називають системою убіквітінірованія). Маркером ж служить ланцюжок не менше ніж з чотирьох молекул білка убіквітину, що складається з 76 амінокислотних залишків. Як освіту ланцюжка через залишок лізину-48 в кожній молекулі, так і приєднання її до білка-субстрату якраз і обслуговується системою ферментів.
Ця система, що включає три типи ферментів (Е1, Е2 і Е3), високо специфічна і вибіркова за рахунок того, що побудована за принципом ієрархічного ускладнення. Фермент Е1 (в клітці він тільки один) активує молекулу убіквітину і передає її одному з ферментів сімейства Е2 (їх називають кон'югується). Потім в каскад реакцій вступає третій учасник - представник сімейства Е3, лигаз, "зшивають" ферментів. Він приймає убіквітин від Е2, з'єднується з білком-субстратом і ковалентно пришиває до нього ланцюжок убіквітину.
Якщо Е1 не має різновидів, то сімейство Е2 налічує 13 членів в клітці дріжджів Saccharomyces cerevisiae, а ссавців - набагато більше. У сімействі Е3 зараз відомо близько 100 різних лигаз, вони-то і визначають в кінцевому рахунку високу специфічність всієї протеолітичної системи. А взагалі кожен фермент і того, і іншого сімейства бере участь в певному клітинному процесі.
Маркування білка-субстрату (мішені) ланцюжком убіквітину завершилася. Тепер її дізнається і зв'язується з нею одна або більше субодиниць регулятора РА700. Цей процес, як і подальше розгортання субстрату, потребує енергії, яку постачають, мабуть,
АТФази базового (нижнього) елемента в структурі того ж регулятора. Розгорнута, лінійна молекула білка простягається через регулятор, який грає роль рота протеасоми, і через відкритий отвір в a -Кільця проникає в протеолитическую камеру. Тут білок розщеплюється на поліпептиди довжиною від 5 до 24 амінокислотних залишків, які вивільняються з протеасоми і в цитоплазмі можуть піддатися гідролізу до амінокислот протеазами (наприклад, ендопептидаз). Непотрібна більше маркировочная ланцюжок ліквідується: ізопептідази розривають її на мономери.
Описуючи каскад реакцій за маркуванням субстрату, ми, щоб не розбивати хід подій, навмисно дещо опустили. Чи не сказали, чому ланцюжок убіквітину пришивається саме до того білку, чия доля вирішена. Виявляється, він уже несе ознаки смерті - специфічні сигнали, які включають процес деградації. Ними можуть бути ділянки всередині білкової молекули або на її N-кінці. Мабуть, в певних умовах вони стають доступними для впізнавання ферментної системою, відповідальною за маркування.
Деякі N-кінцеві амінокислотні залишки (у евкаріот, особливо Арг, Ліз, Лей, Фен, Асп) відіграють велику роль у визначенні життя багатьох короткоживучих білків (в середньому вони існують від декількох хвилин до трьох годин), а також частково зруйнованих або зі зміненою третинної структурою. На залежність швидкості деградації від природи N-кінцевих амінокислот (правило N-кінця) першим звернув увагу наш колишній співвітчизник А.Варшавскій, він же ввів поняття "короткоживучі білки" [ 3 ]. У ряді випадків дестабілізуючі амінокислоти приєднуються до N-кінця довгоживучих білків специфічними ферментами, після чого такі білки швидко руйнуються Протеасома.
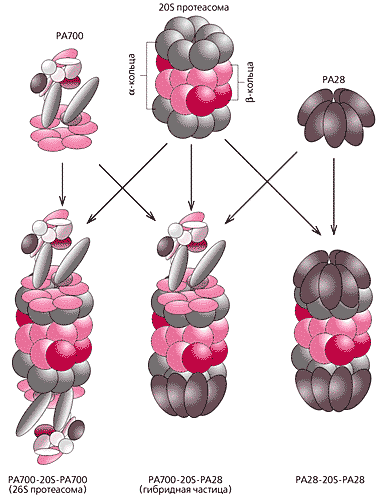
Різні форми протеасоми і її активаторів.
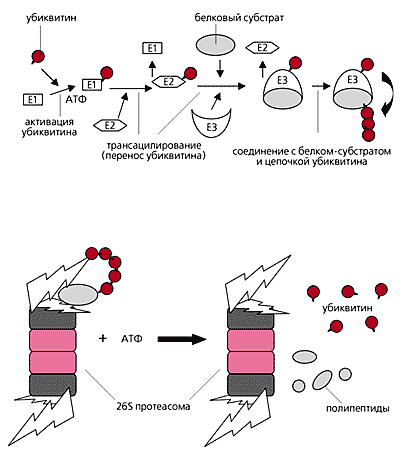
Схема протеасомної деградації білків.
Вгорі - освіту убіквітіновой ланцюжка;
внизу - гідроліз субстрату Протеасома до пептидів і вільного убіквітину.
Ланцюжок убіквітину здатна приєднуватися до білка-мішені і за сигналами, що виникають за рахунок деяких вторинних модифікацій (наприклад, фосфорилювання) або з'єднання з допоміжними білками.
У загальних рисах ми розглянули всю схему (саме схему, тому дуже багато подробиць залишилися за її межами) будови і складання протеасоми, систему маркування білка, який буде нею зруйнований до поліпептидів. За рахунок цього протеасома регулює час життя найважливіших білків, видаляє з неї чужорідні і аномальні, поставляє утворилися в результаті гідролізу поліпептиди в якості антигенів, здатних повідомляти імунній системі про неполадки в клітці. Таким чином, внутрішньоклітинний протеоліз - це не механічний процес деградації білків, а один з основних факторів, які регулюють життєдіяльність клітини.
З різноманітних процесів, в яких бере участь протеасома (табл.), Розглянемо лише деякі.
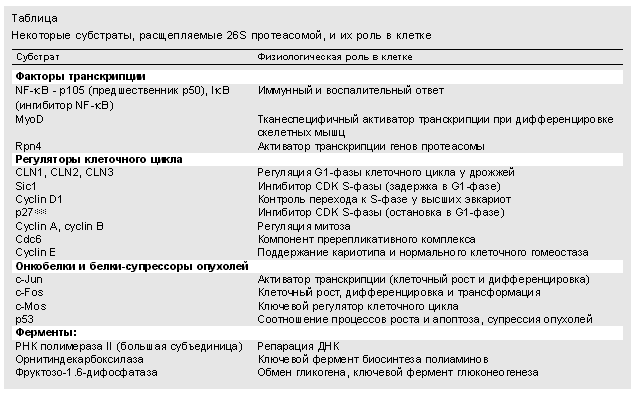
Учасниця багатьох процесів
Відзначимо насамперед, що при вивченні ролі протеасомної деградації в різних клітинних процесах вельми часто використовувалися інгібітори активності протеасоми. Вони легко проникають в клітку і вибірково пригнічують даний шлях протеолізу [4 ].
Регуляція клітинного циклу. Як відомо, клітинний цикл складається з кількох фаз, а їх послідовна зміна регулюється білками цикліни. Оскільки на кожній фазі діє власний регулятор, життя його повинна бути короткою. Це і забезпечує 26S протеасома. Виявилося, що одні цикліни в якості мітки для пізнавання містять ділянки, збагачені пролином, глутамінової кислотою, серином і треонін, а ряд інших - консервативний фрагмент з дев'яти амінокислот, який зазвичай розташовується на відстані близько 40 амінокислотних залишків від N-кінця. Впізнаний по тій чи іншій мітці регуляторний білок зшивається, як уже було сказано, з убіквітіновой ланцюжком своїм власним ферментом з сімейства Е3 і руйнується 26S Протеасома. Ясно, що збій в її роботі викличе зупинку клітинного циклу на тій чи іншій фазі.
Злоякісне переродження клітини. У нормальній здоровій клітині білки, що регулюють швидкість транскрипції, у багатьох випадках визначають її подальшу долю - чи стане працювати і ділитися з запрограмованої для неї швидкістю, чи піде по шляху неконтрольованого злоякісного росту, чи буде зруйнована як представляє небезпеку. Тому такі регуляторні білки в залежності від обставин можуть бути або онкобелкамі, або ж, навпаки, онкосупрессорамі. Приклад подібних перетворень - білок р53, рівнем якого підтримується в здорової клітці співвідношення між процесами росту і апоптозу. Але все змінюється, наприклад, при зараженні людини вірусом папіломи: вірусний білок E6 знаходить білок р53 і сигналізує строго певного ферменту з сімейства Е3 про необхідність приєднати до р53 убіквітіновую ланцюжок. Фермент виконує свою функцію, і р53 стає субстратом для протеасомної деградації. В результаті його прискореного руйнування клітина йде по шляху злоякісного переродження.
Транскрипція. Ми говорили, что під дією 26S протеасоми розщеплюються білкові молекули. Однак з'ясувалося, що протеолизу може піддаватися навіть окрема субодиниця в складному білковому комплексі. Уявити собі механізм взаємодії з ним 26S частки досить важко. Проте частковий протеоліз здійснюється в процесі двоступеневої активації ядерного чинника (його позначають NF-kB), який контролює транскрипцію. Спочатку в результаті гідролізу Протеасома утворюється зі свого попередника субодиниця p50 цього фактора. Потім вона, ще одна субодиниця - р65 - і білок-інгібітор IkB об'єднуються в неактивний комплекс. Транскрипція стає можливою після деградації 26S Протеасома інгібітора IkB, перед цим фосфорилированного в двох місцях.
Імунна система. Участь протеасоми в імунній відповіді клітини - один з найбільш активно досліджуваних аспектів роботи 26S частки. Вона, гидролизуя аномальні або чужорідні білки до поліпептидів, поставляє деякі з них (зазвичай довжиною від восьми до 11 амінокислот) в якості антигенів [ 5 ]. Такі поліпептиди з'єднуються в цитоплазмі з певним транспортним білком і переносяться в ендоплазматичнийретикулум, де взаємодіють з молекулами білків класу I головного комплексу гістосумісності і виносяться на поверхню клітини. Знову з'явилися антигени імунна система виявляє з допомогою цитотоксичних T-лімфоцитів і руйнує клітини, в яких продукуються вірусні або інші незвичайні для них білки.
Щоб ефективніше захищати організм від небезпечних для нього клітин з синтезуються в них аномальними білками, в каталітичному ядрі протеасоми три конституційні b-субодиниці можуть бути замінені на їх ізоформи, або імунні субодиниці. У культурі клітин ссавців така заміна відбувається під дією g-інтерферону - потужного иммуномодулятора. У лімфоїдних органах (селезінці, тимусі, лімфатичних вузлах) ізоформи синтезуються постійно, причому їх кількісне співвідношення з конституційними субодиницями залежить і від типу тканини, і від стадії диференціювання органу. В результаті подібної модифікації кількість утворених Протеасома поліпептидів - потенційних антигенів - зростає в кілька разів.

Але це не єдиний спосіб, за допомогою якого активізується робота протеасоми в якості генератора антигенних поліпептидів.
Ми згадували, що існують три форми протеасоми (в залежності від того, з яким регулятором пов'язано її ядро): РА700-20S-РА700, тобто 26S форма, РА28-20S-РА28 і гібридна РА700-20S-РА28. Припускають, що друга форма здатна гідролізувати довгі пептиди - початковий продукт розщеплення білків 26S Протеасома - до коротких. Саме вони і виносяться на поверхню клітини в якості антигенів. Що стосується гібридної форми, то один її регулятор (РА700), мабуть, дізнається білок, що несе убіквітіновую ланцюжок, а другий (РА28) змушує каталітичне ядро гідролізувати білкову молекулу до більш коротких імуногенних пептидів. До теперішнього часу встановлено, що зміст та внутрішньоклітинний розподіл форм протеасоми пов'язано з дією g-інтерферону і що в цитоплазмі одночасно зі зменшенням кількості 26S протеасоми накопичується гібридна форма.
Термозахист. При нагріванні, як відомо, білки втрачають нормальну конфігурацію і перестають виконувати свої функції. Щоб захистити клітку від настільки згубних наслідків, в ній синтезуються білки теплового шоку, або шаперони. Вони виправляють порушену високою температурою форму білкових ланцюгів. В якій якості бере участь в цьому процесі протеасома, поки не зовсім ясно. Однак встановлено, що, якщо придушити її активність, в клітці накопичуються пошкоджені білки, але в той же час стимулюється синтез мРНК шаперонов. Підвищується також і рівень трегалози (молекули-термозащітніци) і термостабільність клітин в цілому. Отже, вважають, інгібітори протеасоми можуть знайти застосування в медицині як терапевтичних засобів при шокових впливах високими температурами.
Апоптоз. Запрограмована клітинна смерть - один з найважливіших захисних процесів, який забезпечує контроль за кількістю клітин і постійністю складу тканин в організмі. Таким способом руйнуються і небезпечні для нього клітини: заражені вірусами, ракові, з порушеною ДНК. Механізми апоптозу дуже консервативні, однакові у нижчих евкаріот і ссавців і являють собою складну систему послідовних молекулярних подій, які в кінцевому рахунку призводять до ензиматичною фрагментації хромосомної ДНК і смерті клітини. І хоча в апоптозу працюють свої специфічні протеази (каспаз), дослідники виявили, що процес пов'язаний і з протеасомної деградацією білків.
Цю аж ніяк не просту зв'язок вивчають в основному за допомогою інгібіторів активності протеасоми, і найінтенсивніше в дослідженнях онкогенеза. Виявилося, що, знижуючи рівень активності згаданого вже ядерного фактора транскрипції (NF-kB), такі інгібітори можуть запускати апоптоз трансформованих клітин, запобігати ангіогенез (розростання кровоносних капілярів в ракової тканини) і метастазування in vivo. Це результат того, що зниження каталітичної активності протеасоми призводить до накопичення інгібіторів росту клітини і проапоптозних білків. Чи не зруйновані ингибированной Протеасома короткоживучі білки, такі як р53 і р27, теж здатні запустити в клітці каскад біохімічних і морфологічних подій, що ведуть до апоптозу. Однак інгібітори протеасоми можуть і запобігати його, наприклад в первинних клітинних культурах, якщо він викликаний якимись іншими стимулами.
Як видно з наведених прикладів, інгібітори протеасомної деградації служать молекулярним інструментом, за допомогою якого вдається розкрити зв'язок між клітинної смертю і роботою протеасоми. Але не менш важливо й інше: якщо до апоптозу злоякісних клітин призводять інгібітори, можливо, вони можуть послужити медицині як протиракові засоби.
Білки-субстрати і хвороби
Деградація білків Протеасома - процес з тонким налаштуванням, тому збої в ньому, що порушують рівновагу між проліферацією і апоптозом, служать причиною різних хвороб як вроджених, так і набутих. Умовно їх можна розділити на дві групи: захворювання, обумовлені тим, що деградаційна система не працює, і хвороби, які виникають через посилення її функції. Перші - це результат стабілізації субстратів, швидко руйнуються в нормі, другі, навпаки, - аномально швидкого розпаду білків-мішеней.
Ми говорили, що клітинний білок р53 при зараженні онкогенним штамом вірусу папіломи людини з'єднується з вірусним онкобелкі Е6 і стає субстратом для прискореної деградації Протеасома. В результаті позбавлена захисту клітина - мішень вірусної атаки - перетворюється в ракову. Дійсно, при деяких формах карциноми рівень клітинного р53 буває різко знижений. Припускають, що таким способом вірус може неконтрольовано розмножуватися в клітці, і вона стає злоякісною. Агресивні форми раку прямої кишки і молочної залози дослідники теж пов'язують з роботою протеасомної системи, хоча механізм утворення трансформованих клітин тут інший.
Мабуть, деякі вроджені захворювання людини обумовлені генетичними змінами або в білках-субстратах, або в ферментах, що відповідають за зшивання з убіквітин. Фіброкістоз, наприклад (його ознаки - хронічна закупорка і інфекція дихальних шляхів, порушення травлення), пов'язаний з мутацією,
приводить до передчасного гідролізу Протеасома мембранного білка, який регулює транспорт іонів хлору через мембрану епітеліальних клітин. При синдромі Ангельмана (затримка розумового розвитку) виявлені делеції частині Х-хромосоми і пошкоджені молекули ферменту з сімейства Е3. Рідкісна спадкова форма гіпертензії, або синдром Лиддла, обумовлена делецией в генах субодиниць білка, який формує натрієві канали в клітинах епітелію. В результаті змінюється швидкість гідролізу субодиниць Протеасома і, як наслідок, - баланс між іонами калію і натрію (тобто порушується сольовий гомеостаз).
Цілий ряд відомих аутоімунних захворювань може бути викликаний неправильним розрізанням білка Протеасома. Адже вона гідролізує і власні нормальні клітинні білки, і чужорідні. Але з утворюються пептидів тільки чужі стають антигенами, саме по ним цитотоксические лімфоцити дізнаються заражену клітку і знищують її. Свої ж пептиди не викликають Т-клітинної відповіді. Однак, якщо з якої-небудь причини протеасома невірно розщеплює субстрат, то і з власних білків можуть виникати нетипові пептиди, які розпізнаються Т-лімфоцитами як чужорідні. І тоді клітини організму, нічим не заражені, стають мішенню для атаки. Мабуть, з протеасомної системою пов'язані багато імунні захворювання і запальні реакції, але конкретні механізми їх виникнення аж ніяк не однакові.
Цікаво, що віруси можуть піти з-під контролю імунної системи, причому різними способами. Вірус гепатиту В, наприклад, досягає цього за рахунок взаємодії власного білка Х з субодиницями 20S протеасоми і регулятора РА28. У такій комбінації протеолиз пригнічується і білок Х залишається цілим. А раз немає пептидів-антигенів, імунна система не сприймає заражену клітку. Цитомегаловірус людини не охороняє свої білки від гідролізу, а сприяє руйнуванню білкових молекул головного комплексу гістосумісності, які відповідають за транспорт: пептиди-антигени не потрапляють на поверхню зараженої клітини, і та виживає. Інші вірусні білки взаємодіють з протеасомного АТФаза, збільшуючи швидкість гідролізу внутрішньоклітинних білків і приводячи до посиленого розмноження вірусу.
При багатьох нейродегенеративних захворюваннях (в їх числі хвороби Альцгеймера, Паркінсона та ін.) Виявлено сенільні бляшки, різні тіла включення і дегенеративні волокна, збагачені Убіквітин. Однак лише на цій підставі важко зрозуміти, яка роль протеасомної системи в розвитку даних патологій. Правда, в одному випадку вона проявляється наочніше. В нейронах є білок пресенілін-2 (PS2), який пронизує мембрану і бере участь в транспорті попередників амілоїд і подальшому їх перетворенні в амілоїд b -42. Для нормальної роботи PS2 від нього відщеплюється С-кінцевий фрагмент і руйнується Протеасома. Якщо придушити її роботу інгібіторами, фрагмент буде накопичуватися, до такого ж результату приводять мутації в гені білка PS2. А вони виявлені в більшості випадків раннього прояви сімейної форми хвороби Альцгеймера. Таким чином, в її патогенезі може грати роль накопичення С-кінцевого фрагмента, викликане мутаціями в гені або дією інгібіторів протеасоми.
Встановлено, що в дистрофії скелетних м'язів, що зумовлена голодуванням, сепсисом і денервацией, винен посилений гідроліз м'язових білків Протеасома. Але активують її позаклітинні стимули і сигнальні шляхи поки не ясні.
Припускають, що накопичення окислених білків при старінні пов'язано з порушеннями протеасомної деградації. І це не позбавлено підстави.
* * *
Минуло 20 років з відкриття специфічного внутрішньоклітинного протеолізу. За цей час в світі склалося кілька наукових центрів з вивчення протеасоми, її генів і регуляції їх зчитування. Треба сказати, що в лабораторії структури і функції хроматину Інституту молекулярної біології ім. В.А.Енгельгардта РАН виявлена нова сигнальна ланцюг, яка контролює залежний від АТФ гідроліз білків Протеасома [ 6 , 7 ]. Виявилося, у пекарських дріжджів (Saccharomyces cerevisiae) короткоживучий білок Rpn4 служить фактором транскрипції, що регулює роботу більше 500 генів, які кодують більшість субодиниць протеасоми і ферменти, що беруть участь в протеасомної деградації білків. Таким чином, вперше показано, що розщеплення білків контролюється і на рівні транскрипції генів.
Зараз вже ніхто не сумнівається, що без внутрішньоклітинного протеолізу, який специфічно "виводить з гри" в потрібний час і в потрібному місці найважливіші білкові компоненти, клітина не може обійтися. Зроблено дуже багато, а має бути ще більше, тому що робота протеолітичної частки найтіснішим чином замикається з патогенезом багатьох захворювань. А значить, і з можливістю змінювати їх перебіг.
Робота підтримується Російським фондом фундаментальних досліджень. Проект 03-04-49127.
література
1. Wilk S., Orlowski M. // J. Neurochem. 1983. V.40. P.842-849.
2. Arrigo A.-P., Tanaka K., Goldberg AL, Welch WJ // Nature. 1988. V.331. P.192-194.
3. Bachmair A., Finley D., Varshavsky A. // Science. 1986. V.234. P.179-186.
4. Rock KL, Gramm C., Rothstein L. et al. // Cell. 1994. V.78. P.761-771.
5. Rock KL, Goldberg AL. // Ann. Rev. Immunol. 1999. V.17. P.739-777.
6. Mannhaupt G., Schall R., Karpov V. et al. // FEBS Lett. 1999. V.450. P.27-34.
7. Капранов А.В., Курятова А.В., Преображенська О.І., Карпов В.Л. // Мол. біологія. 2001. №35. С.356-364.
8. Baumaister W., Walz J., Zuhl F., Seemuller E. // Cell. 1998. V.92. P.367-380.
Може бути, вона справляється з ними за допомогою добре вивченого протеолізу (розпаду білків під дією спеціальних ферментів)?
Як виникає настільки складна система?
Як же вони проникають в каталітичні області, якщо вхід закритий?

