

67 гостей
бактерії
- бактерії Бакта е рії (грец. Bakterion - паличка), велика група (тип) мікроскопічних, переважно одноклітинних...
- бактерії
- бактерії
- бактерії
- бактерії
- бактерії
бактерії
Бакта е рії (грец. Bakterion - паличка), велика група (тип) мікроскопічних, переважно одноклітинних організмів, що володіють клітинною стінкою, що містять багато дезоксирибонуклеїнової кислоти (ДНК), що мають примітивне ядро, позбавлене видимих хромосом і оболонки, що не містять, як правило, хлорофілу і пластид, що розмножуються поперечним діленням (рідше перетяжкою або брунькуванням). Переважна кількість видів Б. має паличкоподібну форму. Однак до Б. відносять також мікроорганізми, що мають кулясту, ниткоподібну або звиту форму. Б. різноманітні по своїй фізіології, біохімічно дуже активні і поширені в грунті, воді, грунті водоймищ і пр. Б. не уявляють єдиної групи, а виникли різними шляхами. Деякі Б. (наприклад, нитчасті, азотобактер і ін.) Близькі до синезеленим водоростям , Ін. Б. споріднені променистим грибкам - актиноміцетам , Спірохети і деякі ін. Б. мають схожість з одноклітинними тваринами - найпростішими .
Б. беруть участь в круговороті речовин в природі, деякі з них викликають захворювання людини, тварин або рослин, застосовуються в різних галузях мікробіологічної промисловості. Науку, що вивчає Б., називають бактериологией . Це складова частина ширшої дисципліни - мікробіології , В завдання якої входить вивчення всіх сторін життєдіяльності не тільки Б., але і інших мікроорганізмів (дріжджі, цвілеві гриби, мікроскопічні водорості).
Людина використовував Б., ще не знаючи про їхнє існування. За допомогою заквасок, що містять Б., готували кисломолочні продукти, оцет, тісто і т.д. Вперше Б. побачив А. Левенгук - творець мікроскопа, досліджуючи рослинні настої і зубний наліт. До кінця 19 - початку 20 ст. було виділено велике число Б., що мешкають в грунті, воді, харчових продуктах і т.п., були відкриті багато видів хвороботворних Б. Класичні дослідження Л. Пастера в області фізіології Б. послужили основою для вивчення у них обміну речовин. Внесок у дослідження Б. внесли російські і радянські вчені С. Н. Виноградський , В. Л. Омелянський , Б. Л. Ісаченко , Які з'ясували роль Б. в круговороті речовин в природі, який робить можливим життя на Землі. Цей напрямок в мікробіології нерозривно пов'язане з розвитком геології, біогеохімії, ґрунтознавства, з вченням В. І. Вернадського про біосфері .
Морфологія і систематика Б. Розмір, форма, будова, рухливість Б. Діаметр кулястих Б. зазвичай дорівнює 1-2 мкм, ширина паличковидних форм коливається від 0,4 до 0,8 мкм, довжина дорівнює 2-5 мкм. Рідше зустрічаються дуже великі Б. Так, серобактерія Thiophysa macrophysa має 20 мкм в діаметрі, нитки інших серобактерий (Beggiatoa) видно неозброєним оком. Є також дуже дрібні Б., наприклад Bdellovibrio, що паразитують на Б. звичайних розмірів. Деякі Б., наприклад збудники плевропневмонії рогатої худоби, настільки дрібні, що невидимі в оптичний мікроскоп. Кулясті бактерії називаються кокками , Якщо ж вони розташовуються попарно, - диплококами (Рис. 1). Якщо коки розмножуються поперечним діленням і після ділення залишаються з'єднаними, утворюючи ланцюжки, то їх називають стрептококками (Рис. 2, 12). При діленні клітин в трьох взаємно перпендикулярних напрямках утворюються пакети кліток, типові для сарцин (Рис. 3). При розподілі коків в різних площинах виникають скупчення клітин у вигляді грона винограду, що характерно для стафілококів (Рис. 4). Паличкоподібні Б., що утворюють спори, називаються бацилами . Паличкоподібні форми можуть мати «обрубані» або опуклі кінці (рис. 5) і розташовуються окремо або, рідше, у вигляді ланцюжка (рис. 6). Б., що утворюють довгі нитки, - нитчасті бактерії , Мешкають переважно у воді. Б. у формі коми - вібріони (Рис. 7), покручені форми з грубими спіральними завитками - спірили (Рис. 8), з декількома рівномірними тонкими завитками - спірохети (Рис. 9).
Все Б. мають клітинну стінку (рис. 25, 26, 28). Вона виразно видно при приміщенні Б. у розчин кухонної солі; при цьому вміст клітини стискається і відстає від стінки - настає плазмоліз (Рис. 10). У ряду Б. стінка оточена слизової капсулою (рис. 27), присутність якої може бути встановлено при приміщенні таких Б. в розчин туші (рис. 11). При електронній мікроскопії видно, що клітинна стінка складається з декількох шарів (зазвичай трьох). До її складу входять мураміновая кислота, амінокислоти, ліпіди, глюкозамін і інші з'єднання. Хімічний склад клітинної стінки у різних систематичних груп, а також у Б., забарвлюються і не забарвлюються за Грамом, різний. Велику роль в обміні речовин грає цитоплазматична мембрана, яка перебуває під клітинною стінкою. У мембрані (рис. 25, 28) зосереджені численні ферментні системи бактеріальної клітини. У цитоплазмі є рибосоми , До складу яких входить РНК. Вміст нуклеїнових кислот у Б. коливається від 10 до 22% при різному ставленні РНК / ДНК (у кишкової палички воно дорівнює 2). За допомогою електронного мікроскопа встановлено присутність в клітці Б. ниток ДНК, що утворюють ядро, позбавлене оболонки, - т. Зв. нуклеоид (рис. 25, 26, 28). Будова ядра неоднаково у різних Б. Так, у «вищих», складніше організованих Б. (Myxobacteriales, Hyphomycrobiales), ядра легко можуть бути виявлені при мікроскопії забарвлених препаратів в оптичному мікроскопі (рис. 1). У багатьох Б. цитоплазма ущільнена на кінцях кліток і утворює забарвлюються полярні зерна (рис. 13; рис. 2). Клітини Б. містять запасні речовини: жирові включення (рис. 5), зерна глікогену, метахроматіна (рис. 4), гранульози (рис. 6), а також вакуолі, що містять рідину або газ. На відміну від грибів, Б. не містять мітохондрій , Що свідчить про більш примітивному будову Б. Багато Б. рухливі. Зазвичай такі форми мають довгі джгутики, що складаються з скорочувального білка (рис. 14). Завдяки хвилеподібним і спіральним рухам джгутиків клітка Б. переміщається. Види з одним джгутиком на полюсі клітини зв. монотріхі (рис. 15), клітини, що мають пучок джгутиків на кінці, - лофотрихи (рис. 16), Б., у яких джгутики розташовані по всій поверхні тіла, - перітріхі (рис. 17). У міксобактерій , Які також рухливі, джгутиків немає, і вони пересуваються в результаті набухання в довкіллі слизу, що виділяється клітинами (реактивний спосіб руху).
Життєвий цикл. Зміна морфології клітин Б. в часі дає уявлення про їх життєвому циклі. Так, багато аеробних і анаеробних Б. утворюють овальні або круглі блискучі спори . Такі види Б. називаються спороносними (або бацилами). Якщо суперечки великі і розташовуються в центрі клітини, то паличка набуває веретеноподібної форми (рис. 20); у інших видів спору розташовується на кінці палички, і тоді остання набуває форму булави (рис. 19) або барабанної палички. У багатьох спороносних бактерій діаметр спори невеликий, і тому при утворенні спори зберігається паличкоподібна форма Б. (рис. 18, 21). Надалі залишки вегетативної клітини руйнуються, і суперечки стає вільною (рис. 22). У кожній клітині утворюється тільки одна спору і, отже, спорообразование не можна розглядати як розмноження. Спори Б. дуже стійкі до дії високої температури і отруйних речовин. Потрапивши в сприятливе живильне середовище, спори проростають і з них виходять молоді паличкоподібні вегетативні клітини (рис. 23). Цикл розвитку Б. може бути різним. Так, мікобактерії розмножуються як діленням, так і брунькуванням (рис. 25). У міксобактерій вегетативні клітини стискаються, скорочуються і утворюють круглі або овальні мікроцисти (див. циста ), Які потім можуть проростати (рис. 24). Сполучені слизом мікроцисти утворюють тіла кулястої, грибовидной або Коралоподібних форми зеленого, рожевого або іншого кольору. В процесі зростання Б. можуть утворювати фільтрівні форми, що проходять через фільтри і дають в подальшому культури, подібні або тотожні з тими, в яких вони виникли.
Мінливість, фізіологічні і морфологічні властивості Б. можуть піддаватися змінам. Б. можуть втратити рухливість, здатність утворювати пігменти, давати суперечки, посилювати або зменшувати здатність до синтезу різних органічних сполук, змінювати форму і будову колоній на щільних поживних середовищах і т.д. Ці зміни можуть відбуватися мимовільно, т. Е. Безпрімененія відповідних зовнішніх впливів. Значно більше змінених форм виникає в результаті застосування мутагенів (ультрафіолетові промені, іонізуюча радіація, етиленімін або інших хімічних речовини). Кожна властивість Б. пов'язано з ДНК, т. Е. Контролюється відповідним геном . завдяки успіхам генетики мікроорганізмів встановлено місце розташування багатьох генів в нитки ДНК (див. Генетичні карти хромосом ). Виділивши ДНК з клітин-мутантів і додавши її в культуру іншого штаму, можна викликати (в результаті проникнення ДНК всередину клітин) спадкові зміни, називаються трансформацією . За допомогою мутагенів можуть бути отримані мутанти, цінні в практичному відношенні, в тому числі утворюють більшу кількість різних антибіотиків, амінокислот, вітамінів та інших біологічно активних речовин. За допомогою мутантів Б. були розшифровані шляху біосинтезу різних органічних сполук. Поступово змінюючи середовище проживання Б., можна адаптувати їх до нових умов існування. Так були отримані форми, стійкі до різних отрут, що розвиваються при незвичайній температурі або реакції середовища, і т.п .; так з'являються стійкі до деяких антибіотиків форми хвороботворних Б.
Систематика. Для з'ясування систематич. положення Б. визначають їх розміри, морфологію клітин, характер зростання чистої культури на різних питати. середовищах, форму, колір і характер поверхні колоній, що виростають на щільних середовищах. Встановлюють також характер розрідження Б. желатин (рис. 30), здатність їх згортати молоко, зброджувати різні вуглеводи, відновлювати нітрати, ооразовивать аміак, сірководень і індол при розкладанні білків і т.п. Надалі, маючи характеристику виділеної культури, визначають її систематичне положення. Б. підрозділяють на три класи. Перший клас - Eubacteria - об'єднує Б., що мають щільну клітинну стінку і не утворюють плодових тіл. В цьому класі розрізняють наступні порядки: 1) Eubacteriales - одноклітинні коки, неветвящиеся палички і спірально звиті форми; до цього порядку відносяться всі неспороносні і спороносні Б., фото-синтезують Б., спірохети та ін .; 2) Trichobacteriales - багатоклітинні нитчасті Б. з поперечними перегородками; 3) Ferribacteriales - одноклітинних нитчасті автотрофні железобактерии; 4) Thiobacteriales - одноклітинні автотрофні серобактерии. Другий клас - Myxobacteria - об'єднує Б. з тонкою клітинною стінкою і реактивним характером руху, що утворюють мікроцисти і плодові тіла різної форми. До третього класу - Hyphomicrobiales - відносять клітки, що дають довгі нитки, на кінцях яких утворюються нирки; відокремилися нирки рухливі. Сов. систематики виділяють на підставі еволюційних (філогенезі) даних великі групи, які об'єднують родинні форми. Так, наприклад, розгалужені мікобактерії не виділяються в самостійну групу, а об'єднані з актиноміцетами.
Фізіологія Б. Зростання, розмноження, розвиток. Після поділу Б. кожна з двох дочірніх бактеріальних клітин починає рости і досягає розмірів материнської. У цьому випадку говорять про зростання окремої клітини. Розмноження клітин, складових популяцію , Призводить до збільшення загального числа клітин. У цьому випадку говорять про зростання культури. При зростанні культури в рідкому поживному середовищі остання стає каламутною; чим більше клітин в культурі, тим вона мутніше. Про інтенсивність росту судять на підставі підрахунку клітин в 1 мл культури за допомогою мікроскопа або визначають за допомогою нефелометра ступінь каламутності живильного середовища. Визначаючи кількість клітин в різні періоди зростання культури, можна отримати криву зростання, що відображає кілька фаз: спочатку клітини не розмножуються, потім починають ділитися, причому швидкість розмноження весь час зростає; далі настає фаза, для якої характерна постійна швидкість ділення клітин; потім ця швидкість зменшується і настає відмирання клітин. Для отримання максимальної кількості клітин Б. вирощують в умовах т. Н. проточною культури; при цьому з посудини, в якому розмножуються Б., витікає певний обсяг культури; одночасно в судину додається в такій же кількості свіжа стерильна живильне середовище. При розмноженні Б. не в проточних, а в стаціонарних умовах відбувається зміна живильного середовища і накопичення в ній продуктів життєдіяльності Б., унаслідок чого змінюються і їх фізіологічні особливості. Так, молоді клітини Clostridium acetobutylicum не здатні утворювати ацетон; це властивість вони набувають в старішої культурі. Якщо спороносних Б. вирощувати в умови проточної культури, вони ділитимуться, але не даватимуть суперечку. При вирощуванні Б. на щільних поживних середовищах вони утворюють скупчення клітин різних розмірів, форми, кольору, звані колоніями.
Харчування. До складу клітин Б. входять ті ж біогенні елементи і мікроелементи, що і до складу клітин вищих рослин і тварин. Це С, N, О, Н, S, Р, К, Mg, Ca, Cl, Fe н ін. Крім білка, вуглеводів і жирів, Б. містять також РНК і велика кількість ДНК. Всі ці речовини можуть бути синтезовані тільки з речовин, що містяться в навколишньому середовищі. Як правило, через напівпроникну клітинну стінку і цитоплазматичну мембрану всередину Б. проходять тільки розчинні речовини. Під дією гідролітичних ферментів, що надходять з бактеріальних клітин назовні, відбувається розкладання складніших речовин (наприклад, крохмалю, целюлози) з утворенням розчинних продуктів (наприклад, моносахаров), засвоюваних Б. Як джерело азоту Б. можуть засвоювати білки, амінокислоти, амонійні солі , нітрати. Різні види Б. здатні утилізувати різні джерела азоту. Раніше вважали, що деякі патогенні (хвороботворні) і молочнокислі Б. можуть розвиватися лише в живильних середовищах з білками. Надалі з'ясувалося, що джерелом азоту для таких Б. можуть служити амонійні солі. Існує багато видів Б. з різних систематичних груп, які здатні засвоювати не тільки азот тих чи інших азотовмісних речовин, але і фіксувати азот атмосфери. До таких азотфиксирующим мікроорганізмам відносяться азотобактерии, мікобактерії, пурпурні фотосинтезирующие Б., а також бульбочкові бактерії (Див. Азотфіксация ). Джерелами мінерального живлення для Б. служать солі Р, S, Cl, К, Fe, Na, Ca; багато видів потребують також мікроелементів (Mo, Мn, Си, В, V і ін.). Для розмноження Б. необхідні також чинники зростання мікроорганізмів, до яких належать вітаміни групи В, біотин, пантотенова кислота, фолієва кислота та ін. Б., здатні синтезувати ці речовини, називаються ауксоавтотрофамі. До них відносяться псевдомоноси і багато інших неспороносні Б. При вирощуванні Б., не здатних до синтезу чинників зростання, їх додають в живильне середовище. Такі Б. називаються а у ксогетеротрофам і, до них відносяться, наприклад, різні молочнокислі Б. Джерелами вуглецю для Б. можуть служити вуглеводи, спирти, органічні кислоти, лігнін, хітин, вуглеводні, жири та ін. Здатність засвоювати вуглець з тих чи інших джерел у різних видів Б. варіює, і цим користуються для цілей систематики. Б., що засвоюють вуглець органічних сполук, називаються гетеротрофних. Б., що засвоюють вуглець вуглекислого газу атмосфери, називаються автотрофними. Види, що використовують для фіксації вуглекислоти енергію сонячних променів, наз. фотоавтотрофамі. Група Б., одержуючих енергію в результаті окислення таких неорганічних речовин, як аміак, нітрити, сірка, водень і ін., Здатних засвоювати вуглекислоту за рахунок енергії, що звільняється при окисленні вказаних неорганічних з'єднань, називаються хемоавтотрофов і, а сам процес асиміляції двоокису вуглецю, відкритий видатним російським мікробіологом С. Н. Виноградским, - хемосинтезом .
Дихання и обмін Речовини. Синтез Речовини, что входять до складу бактеріальної Клітини, ее рухлівість и інші процеси супроводжуються витрати ЕНЕРГІЇ. Більшість Б. отрімує Енергію Шляхом окислення органічніх Речовини, хемоавтотрофні - в результате окислення неорганічніх Сполука, фотосинтезирующие Б. Використовують Енергію Сонячних променів. Б., здатні рости только в прісутності кисня, назіваються аеробамі , Що ростуть во время відсутності кисня, - анаероби . При аеробних діханні відбувається окислення органічніх Сполука з віділенням вуглекислий газу. Если ж окислення идет не до кінця, то в середовіщі накопічуються проміжні продукти. Такі процеси назіваються окіслювальнімі бродіння (например, уксуснокислое бродіння). Розкладання органічніх Речовини в анаеробних условиях Із звільненням ЕНЕРГІЇ назіваються бродінням . При зброджуванні вуглеводів різнімі Б. могут утворюватіся: молочна або масляна кислота, етиловий, пропіловій або бутиловий спирт, ацетон и інші Речовини. Ряд біохімічніх процесів (гліколіз, транспорт електронів, цикл Кребса, синтез амінокіслот, білків, нуклеїнових кислот и ін.) Протікає у Б. почти так само, як в клітінах рослин и тварин. Специфічне особливості обміну речовин Б. - висока біохімічна активність, здатність окислювати неорганічні сполуки сірки, азоту (аміак) і ін., Синтезувати білок, використовуючи в якості вихідного продукту фенол, метан та інші вуглеводні, окисляти водень, фіксувати азот атмосфери, синтезувати ферменти, розкладають целюлозу або лігнін, утворювати метан з вуглекислоти і водню та ін. - виключно цінні в практичному відношенні.
Екологія Б. Поширення. Б. відносяться до космополітів. Одні і ті ж види Б. можна знайти на всіх материках, т. Е. Майже повсюдно. Кількість їх в грунті, воді та інших середовищах визначають прямим підрахунком клітин в пофарбованому препараті або посівом на різні поживні середовища. В 1 г грунту містяться сотні тисяч або мільйони бактерій; в 1 мл води - десятки або сотні клітин. Сильний вплив на бактеріальну мікрофлору надають екологічні умови. Так, окультурені грунту не тільки містять більше Б., чим, наприклад, грунти пустель, але і розрізняються за видовим складом мікрофлори. Сучасній мікробіології відомо не більше 1/10 частини Б., що існують в природі. Застосування методів капілярної і електронної мікроскопії препаратів грунту дозволило виявити багато нових видів Б. Розвиваючись в самих різних екологічних умовах, Б. в процесі еволюції пристосувалися до них. Так виникли термофільні Б., що мешкають у воді гарячих джерел, в розігрівають купах торфу або гною, психрофільні форми, що живуть при низькій температурі в воді полярних морів, галофільні Б., здатні розмножуватися в середовищі, що містить до 20% кухонної солі, ацидофільні та алкалофільних Б., зростаючі в дуже кислою або сильно лужному середовищі, і т.д. Широке поширення в природі певних джерел вуглецю або азоту привело в ході еволюції до фізіологічної конвергенції , Т. Е. Появі у представників різних систематичних груп Б. здатності засвоювати біогенний елемент з одного джерела. Так, Б., здатні фіксувати атмосферний азот, належать до різних класів, порядків і сімейств; А; здатністю утилізувати целюлозу мають багато Б., далекі в систематичному відношенні. Між різними видами. Б., з одного боку, і іншими мікроорганізмами, рослинами або тваринами - з іншого, можуть існувати як антагоністичні (див. антагонізм ), Так і симбіотичні (див. симбіоз ) Відносини. Деякі Б. утворюють пігменти, антибіотики або органічні кислоти, що пригнічують життєдіяльність інших Б., грибів, водоростей, одноклітинних і деяких клітин багатоклітинних тварин. Бактеріальні віруси - бактеріофаги - проникають всередину Б. і, розмножуючись в них, викликають загибель і лізис мікроорганізмів. При симбіотичних, т. Е. Заснованих на взаємній користі, стосунках один вид Б. може споживати продукти життєдіяльності іншого виду, накопичення яких в культуральній рідині гальмує зростання останнього. У свою чергу симбіонт може виділяти в середу додаткові чинники зростання, необхідні іншого виду. Б., що мешкають в кишечнику тварин або людини і харчуються за рахунок вмісту кишечника, утворюють ферменти, необхідні для травлення, а також ряд речовин, вкрай важливих для життя господаря (незамінні амінокислоти, різні вітаміни та ін.).
Роль Б. в круговороті речовин в природі. Мінералізуя рослинні і тваринні залишки, мікроорганізми беруть участь у кругообігу всіх хімічних елементів, що входять до складу живих клітин. Так, джерелом вуглецю для вищих рослин і хемоавтотрофні Б. служить вуглекислота атмосфери, що фіксується в процесі фото- або хемосинтезу. біомаса рослин і тварин розкладається мікроорганізмами, здатними утилізувати целюлозу, пентози, крохмаль, лігнін, пектинові речовини, в кінцевому підсумку до вуглекислоти і води (рис. 31).
Так само велика роль мікроорганізмів, в тому числі Б., в кругообігу азоту. Тварини, харчуючись рослинами, синтезують білок та інші азотовмісні продукти свого тіла за рахунок білка рослин. При мінералізації тваринного і рослинного білка гнильні Б. утворюють аміак, який окислюється нітріфіцірующімі Б. в нітрити і потім в нітрати. Як амонійні солі, так і нітрати служать джерелом азотистого живлення для вищих рослин, що синтезують при цьому білки свого тіла (рис. 32). Мінераліз здатність Б. забезпечує круговорот і інших біогенних елементів. Руйнуючи органічні сполуки фосфору (нуклеїнові кислоти і ін.), Вони збагачують мінеральними з'єднаннями фосфору водоймища і грунт. Під впливом Б. відбувається мінералізація і органічних сполук сірки. Сірчані Б. можуть окислювати сірководень, сірку або деякі її сполуки до сірчаної кислоти, інші здатні відновлювати сульфати з утворенням сірководню. Б. здійснюють окислення заліза і марганцю, відкладення солей кальцію, окислення метану і водню, руйнування гірських порід продуктами життєдіяльності і ін. Все це дозволяє вважати Б. потужними геологічними діячами.
Практичне значення. Б. служать улюбленими об'єктами для вирішення загальних питань генетики, біохімії, біофізики, космічної біології і ін. Культури Б. застосовуються для кількісного визначення амінокислот, вітамінів, антибіотиків. Родючість ґрунтів значною мірою пов'язано з життєдіяльністю Б., минерализующих рослинні і тваринні залишки з утворенням з'єднань, що засвоюються с.-г. рослинами. Разом з тим, синтезуючи живу речовину клітин, Б. накопичують великі кількості органічних сполук в грунті. У верхніх шарах окультуреній грунту на площі в 1 га міститься декілька т бактеріальних клітин. Живучи в грунті азотфіксуючі Б. збагачують грунт азотом. Виключно велика роль бульбочкових бактерій, які фіксують газоподібний азот. Зараження насіння бобових рослин нитрагином - препаратом, що містить клітини бульбочкових Б. (див. бактеріальні добрива ), Підвищує урожай рослин і накопичення азоту в грунті. За допомогою Б., що зброджують пектинові речовини, здійснюють мочку льону, конопель, кенафа та інших луб'яних культур. Різні види Б. застосовують при отриманні з молока кисломолочних продуктів, масла і сиру.
У мікробіологічної промисловості за допомогою соответстветствующих видів Б. отримують з крахмалсодержащего або іншої сировини молочну кислоту, ацетон, етиловий, бутиловий та інші спирти, кровозамінник декстран, діацетил, антибіотики (граміцидин і ін.), Вітаміни, амінокислоти та ін. Особливо широко застосовуються Б . для отримання ферментних препаратів (амілази, протеази і ін.). В результаті розмноження Б., створюючих молочну кислоту з вуглеводів, квашена капуста, силос, солоні огірки не гниють, тому що кисла реакція заважає розвитку гнильних Б. Окисляющие сірку Б. застосовують для бактеріального вилуговування міді і ряду інших металів з містять їх порід. Помістивши здатні засвоювати газоооразние вуглеводні Б. у судини, що зариваються потім в грунт, можна на підставі зростання Б. укладати, є чи в цій місцевості нафта або природний газ. З багатьма видами Б. доводиться вести серйозну боротьбу, оберігаючи від псування і руйнування ними зерно, овочі, фрукти, всі харчові продукти, різні види сировини, матеріалів і виробів (текстиль, картон, канати, риболовні сіті, ізоляція кабелю і мн. Ін. ). Багато хвороб людини викликаються патогенними Б. До таких хвороб відносяться різні епідемічного захворювання (холера, черевний тиф, паратифи, чума, дифтерія, туляремія, бруцельоз), а також туберкульоз, зараження крові (сепсис), проказа, сифіліс та ін. У тварин Б . викликають сап, сибірську виразку, туберкульоз і ін. Багато хвороб як культурних, так і диких рослин викликають т. н. фітопатогенні Б. (див. Бактеріальні хвороби рослин ). Боротьба з хвороботворними Б. грунтується на асептики и антисептику , На застосуванні бактериостатических речовин и бактерицидних речовин (Див. антибіотики , хіміотерапія ).
Літ .: Красильников Н. А., Визначник бактерій і актиноміцетів, М.-Л., 1949; Ісаченко Б. Л., Вибрані праці, т. 1-2, М.-Л., 1951; Виноградський С. Н., Мікробіологія грунту, М., 1952; Кузнєцов С. І., Роль мікроорганізмів у кругообігу речовин в озерах, М., 1952; Імшенецький А. А., Мікробіологія целюлози, М., 1953; Омелянський В. Л., Вибрані праці, т. 1-2, М., 1953; Анатомія бактерій, пров. з англ., М., 1960; Работнова І. Л., Загальна мікробіологія, М., 1966; Clifton С. Є., Introduction to bacterial physiology, NY, 1957; Gunsalus 1. С. and Stanier RJ, The Bacteria, v. 1-5, NY, 1960; Stanier RJ, Dondoroff M., Adelberg Е. A., Themic-robial world, 2 ed., NY, 1963; Lamanna C., Mallette М. F., Basic bacteriology, 3 ed., Baltimore, 1965.
А. А. Імшенецький.
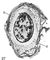
Мал. 27 до ст. Бактерії. Слизова капсула (а) у цисти азотобактера.
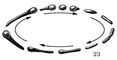
Мал. 23 до ст. Бактерії.

Включення запасного речовини - гранульози (забарвлюється йодом в синій колір) в клітинах анаеробної бактерії під час утворення нею суперечка.

Мал. 9 до ст. Бактерії. Спірохети.

Мал. 4 до ст. Бактерії. Стафілококи.

Мал. 30 до ст. Бактерії.

Мал. 25 до ст. Бактерії. Клітка золотистого стафілокока: а - клітинна стінка; б - цитоплазматична мембрана; в - нуклеоїд; г - мембранні структури всередині нуклеоида.
Мал. 2 до ст. Бактерії. Стрептококи.

Мал. 11 до ст. Бактерії.

Мал. 18 до ст. Бактерії.

Мал. 28 до ст. Бактерії. Клітка молочнокислої бактерії; а - внутрицитоплазматические мембранні структури (мезосоми); б - клітинна стінка; в - цитоплазматична мембрана.

Гігантські форми бактерій, що виникають при несприятливих умовах життя.

Мал. 19 до ст. Бактерії.

Клітини оцтовокислих бактерій із зернами метахроматіна.

Мал. 3 до ст. Бактерії. Сарціни.

Мал. 8 до ст. Бактерії. Спірілли.

Полярні зерна ( «шапочки»); чітко фарбуються у багатьох спороносних бактерій.

Мал. 15 до ст. Бактерії.

Мал. 17 до ст. Бактерії.

Мал. 13 до ст. Бактерії.

Клітини спороносной бактерії з великими краплями жиру.
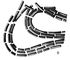
Мал. 6 до ст. Бактерії. Бацили.

Мал. 20 до ст. Бактерії.

Мал. 29 до ст. Бактерії. Джгутик кишкової палички.

Мал. 26 до ст. Бактерії. Освіта поперечної перегородки в делящейся клітці гемолітичного стрептокока: а - утворюється перегородка; б - нуклеоїд; в - клітинна стінка.

Мал. 22 до ст. Бактерії.

Мал. 10 до ст. Бактерії. Плазмоліз.

Ядра в клітинах миксобактерии Polyangium; клітини розмножуються перетяжкою.

Мал. 1 до ст. Бактерії. Диплококи.
Мал. 32 до ст. Бактерії.

Мал. 12 до ст. Бактерії. Стрептококи.
Метахроматін в клітинах збудника дифтерії.

Мал. 5 до ст. Бактерії. Бацили.

Мал. 21 до ст. Бактерії.

Великі вигнуті клітини (спірили) із зернами метахроматіна.
Мал. 31 до ст. Бактерії.
Мал. 7 до ст. Бактерії. Вібріони.

Мал. 16 до ст. Бактерії.

Кислотостійкі палички прокази.
Мал. 24 до ст. Бактерії.

Мал. 14 до ст. Бактерії.
бактерії
Бакта е рії (грец. Bakterion - паличка), велика група (тип) мікроскопічних, переважно одноклітинних організмів, що володіють клітинною стінкою, що містять багато дезоксирибонуклеїнової кислоти (ДНК), що мають примітивне ядро, позбавлене видимих хромосом і оболонки, що не містять, як правило, хлорофілу і пластид, що розмножуються поперечним діленням (рідше перетяжкою або брунькуванням). Переважна кількість видів Б. має паличкоподібну форму. Однак до Б. відносять також мікроорганізми, що мають кулясту, ниткоподібну або звиту форму. Б. різноманітні по своїй фізіології, біохімічно дуже активні і поширені в грунті, воді, грунті водоймищ і пр. Б. не уявляють єдиної групи, а виникли різними шляхами. Деякі Б. (наприклад, нитчасті, азотобактер і ін.) Близькі до синезеленим водоростям , Ін. Б. споріднені променистим грибкам - актиноміцетам , Спірохети і деякі ін. Б. мають схожість з одноклітинними тваринами - найпростішими .
Б. беруть участь в круговороті речовин в природі, деякі з них викликають захворювання людини, тварин або рослин, застосовуються в різних галузях мікробіологічної промисловості. Науку, що вивчає Б., називають бактериологией . Це складова частина ширшої дисципліни - мікробіології , В завдання якої входить вивчення всіх сторін життєдіяльності не тільки Б., але і інших мікроорганізмів (дріжджі, цвілеві гриби, мікроскопічні водорості).
Людина використовував Б., ще не знаючи про їхнє існування. За допомогою заквасок, що містять Б., готували кисломолочні продукти, оцет, тісто і т.д. Вперше Б. побачив А. Левенгук - творець мікроскопа, досліджуючи рослинні настої і зубний наліт. До кінця 19 - початку 20 ст. було виділено велике число Б., що мешкають в грунті, воді, харчових продуктах і т.п., були відкриті багато видів хвороботворних Б. Класичні дослідження Л. Пастера в області фізіології Б. послужили основою для вивчення у них обміну речовин. Внесок у дослідження Б. внесли російські і радянські вчені С. Н. Виноградський , В. Л. Омелянський , Б. Л. Ісаченко , Які з'ясували роль Б. в круговороті речовин в природі, який робить можливим життя на Землі. Цей напрямок в мікробіології нерозривно пов'язане з розвитком геології, біогеохімії, ґрунтознавства, з вченням В. І. Вернадського про біосфері .
Морфологія і систематика Б. Розмір, форма, будова, рухливість Б. Діаметр кулястих Б. зазвичай дорівнює 1-2 мкм, ширина паличковидних форм коливається від 0,4 до 0,8 мкм, довжина дорівнює 2-5 мкм. Рідше зустрічаються дуже великі Б. Так, серобактерія Thiophysa macrophysa має 20 мкм в діаметрі, нитки інших серобактерий (Beggiatoa) видно неозброєним оком. Є також дуже дрібні Б., наприклад Bdellovibrio, що паразитують на Б. звичайних розмірів. Деякі Б., наприклад збудники плевропневмонії рогатої худоби, настільки дрібні, що невидимі в оптичний мікроскоп. Кулясті бактерії називаються кокками , Якщо ж вони розташовуються попарно, - диплококами (Рис. 1). Якщо коки розмножуються поперечним діленням і після ділення залишаються з'єднаними, утворюючи ланцюжки, то їх називають стрептококками (Рис. 2, 12). При діленні клітин в трьох взаємно перпендикулярних напрямках утворюються пакети кліток, типові для сарцин (Рис. 3). При розподілі коків в різних площинах виникають скупчення клітин у вигляді грона винограду, що характерно для стафілококів (Рис. 4). Паличкоподібні Б., що утворюють спори, називаються бацилами . Паличкоподібні форми можуть мати «обрубані» або опуклі кінці (рис. 5) і розташовуються окремо або, рідше, у вигляді ланцюжка (рис. 6). Б., що утворюють довгі нитки, - нитчасті бактерії , Мешкають переважно у воді. Б. у формі коми - вібріони (Рис. 7), покручені форми з грубими спіральними завитками - спірили (Рис. 8), з декількома рівномірними тонкими завитками - спірохети (Рис. 9).
Все Б. мають клітинну стінку (рис. 25, 26, 28). Вона виразно видно при приміщенні Б. у розчин кухонної солі; при цьому вміст клітини стискається і відстає від стінки - настає плазмоліз (Рис. 10). У ряду Б. стінка оточена слизової капсулою (рис. 27), присутність якої може бути встановлено при приміщенні таких Б. в розчин туші (рис. 11). При електронній мікроскопії видно, що клітинна стінка складається з декількох шарів (зазвичай трьох). До її складу входять мураміновая кислота, амінокислоти, ліпіди, глюкозамін і інші з'єднання. Хімічний склад клітинної стінки у різних систематичних груп, а також у Б., забарвлюються і не забарвлюються за Грамом, різний. Велику роль в обміні речовин грає цитоплазматична мембрана, яка перебуває під клітинною стінкою. У мембрані (рис. 25, 28) зосереджені численні ферментні системи бактеріальної клітини. У цитоплазмі є рибосоми , До складу яких входить РНК. Вміст нуклеїнових кислот у Б. коливається від 10 до 22% при різному ставленні РНК / ДНК (у кишкової палички воно дорівнює 2). За допомогою електронного мікроскопа встановлено присутність в клітці Б. ниток ДНК, що утворюють ядро, позбавлене оболонки, - т. Зв. нуклеоид (рис. 25, 26, 28). Будова ядра неоднаково у різних Б. Так, у «вищих», складніше організованих Б. (Myxobacteriales, Hyphomycrobiales), ядра легко можуть бути виявлені при мікроскопії забарвлених препаратів в оптичному мікроскопі (рис. 1). У багатьох Б. цитоплазма ущільнена на кінцях кліток і утворює забарвлюються полярні зерна (рис. 13; рис. 2). Клітини Б. містять запасні речовини: жирові включення (рис. 5), зерна глікогену, метахроматіна (рис. 4), гранульози (рис. 6), а також вакуолі, що містять рідину або газ. На відміну від грибів, Б. не містять мітохондрій , Що свідчить про більш примітивному будову Б. Багато Б. рухливі. Зазвичай такі форми мають довгі джгутики, що складаються з скорочувального білка (рис. 14). Завдяки хвилеподібним і спіральним рухам джгутиків клітка Б. переміщається. Види з одним джгутиком на полюсі клітини зв. монотріхі (рис. 15), клітини, що мають пучок джгутиків на кінці, - лофотрихи (рис. 16), Б., у яких джгутики розташовані по всій поверхні тіла, - перітріхі (рис. 17). У міксобактерій , Які також рухливі, джгутиків немає, і вони пересуваються в результаті набухання в довкіллі слизу, що виділяється клітинами (реактивний спосіб руху).
Життєвий цикл. Зміна морфології клітин Б. в часі дає уявлення про їх життєвому циклі. Так, багато аеробних і анаеробних Б. утворюють овальні або круглі блискучі спори . Такі види Б. називаються спороносними (або бацилами). Якщо суперечки великі і розташовуються в центрі клітини, то паличка набуває веретеноподібної форми (рис. 20); у інших видів спору розташовується на кінці палички, і тоді остання набуває форму булави (рис. 19) або барабанної палички. У багатьох спороносних бактерій діаметр спори невеликий, і тому при утворенні спори зберігається паличкоподібна форма Б. (рис. 18, 21). Надалі залишки вегетативної клітини руйнуються, і суперечки стає вільною (рис. 22). У кожній клітині утворюється тільки одна спору і, отже, спорообразование не можна розглядати як розмноження. Спори Б. дуже стійкі до дії високої температури і отруйних речовин. Потрапивши в сприятливе живильне середовище, спори проростають і з них виходять молоді паличкоподібні вегетативні клітини (рис. 23). Цикл розвитку Б. може бути різним. Так, мікобактерії розмножуються як діленням, так і брунькуванням (рис. 25). У міксобактерій вегетативні клітини стискаються, скорочуються і утворюють круглі або овальні мікроцисти (див. циста ), Які потім можуть проростати (рис. 24). Сполучені слизом мікроцисти утворюють тіла кулястої, грибовидной або Коралоподібних форми зеленого, рожевого або іншого кольору. В процесі зростання Б. можуть утворювати фільтрівні форми, що проходять через фільтри і дають в подальшому культури, подібні або тотожні з тими, в яких вони виникли.
Мінливість, фізіологічні і морфологічні властивості Б. можуть піддаватися змінам. Б. можуть втратити рухливість, здатність утворювати пігменти, давати суперечки, посилювати або зменшувати здатність до синтезу різних органічних сполук, змінювати форму і будову колоній на щільних поживних середовищах і т.д. Ці зміни можуть відбуватися мимовільно, т. Е. Безпрімененія відповідних зовнішніх впливів. Значно більше змінених форм виникає в результаті застосування мутагенів (ультрафіолетові промені, іонізуюча радіація, етиленімін або інших хімічних речовини). Кожна властивість Б. пов'язано з ДНК, т. Е. Контролюється відповідним геном . завдяки успіхам генетики мікроорганізмів встановлено місце розташування багатьох генів в нитки ДНК (див. Генетичні карти хромосом ). Виділивши ДНК з клітин-мутантів і додавши її в культуру іншого штаму, можна викликати (в результаті проникнення ДНК всередину клітин) спадкові зміни, називаються трансформацією . За допомогою мутагенів можуть бути отримані мутанти, цінні в практичному відношенні, в тому числі утворюють більшу кількість різних антибіотиків, амінокислот, вітамінів та інших біологічно активних речовин. За допомогою мутантів Б. були розшифровані шляху біосинтезу різних органічних сполук. Поступово змінюючи середовище проживання Б., можна адаптувати їх до нових умов існування. Так були отримані форми, стійкі до різних отрут, що розвиваються при незвичайній температурі або реакції середовища, і т.п .; так з'являються стійкі до деяких антибіотиків форми хвороботворних Б.
Систематика. Для з'ясування систематич. положення Б. визначають їх розміри, морфологію клітин, характер зростання чистої культури на різних питати. середовищах, форму, колір і характер поверхні колоній, що виростають на щільних середовищах. Встановлюють також характер розрідження Б. желатин (рис. 30), здатність їх згортати молоко, зброджувати різні вуглеводи, відновлювати нітрати, ооразовивать аміак, сірководень і індол при розкладанні білків і т.п. Надалі, маючи характеристику виділеної культури, визначають її систематичне положення. Б. підрозділяють на три класи. Перший клас - Eubacteria - об'єднує Б., що мають щільну клітинну стінку і не утворюють плодових тіл. В цьому класі розрізняють наступні порядки: 1) Eubacteriales - одноклітинні коки, неветвящиеся палички і спірально звиті форми; до цього порядку відносяться всі неспороносні і спороносні Б., фото-синтезують Б., спірохети та ін .; 2) Trichobacteriales - багатоклітинні нитчасті Б. з поперечними перегородками; 3) Ferribacteriales - одноклітинних нитчасті автотрофні железобактерии; 4) Thiobacteriales - одноклітинні автотрофні серобактерии. Другий клас - Myxobacteria - об'єднує Б. з тонкою клітинною стінкою і реактивним характером руху, що утворюють мікроцисти і плодові тіла різної форми. До третього класу - Hyphomicrobiales - відносять клітки, що дають довгі нитки, на кінцях яких утворюються нирки; відокремилися нирки рухливі. Сов. систематики виділяють на підставі еволюційних (філогенезі) даних великі групи, які об'єднують родинні форми. Так, наприклад, розгалужені мікобактерії не виділяються в самостійну групу, а об'єднані з актиноміцетами.
Фізіологія Б. Зростання, розмноження, розвиток. Після поділу Б. кожна з двох дочірніх бактеріальних клітин починає рости і досягає розмірів материнської. У цьому випадку говорять про зростання окремої клітини. Розмноження клітин, складових популяцію , Призводить до збільшення загального числа клітин. У цьому випадку говорять про зростання культури. При зростанні культури в рідкому поживному середовищі остання стає каламутною; чим більше клітин в культурі, тим вона мутніше. Про інтенсивність росту судять на підставі підрахунку клітин в 1 мл культури за допомогою мікроскопа або визначають за допомогою нефелометра ступінь каламутності живильного середовища. Визначаючи кількість клітин в різні періоди зростання культури, можна отримати криву зростання, що відображає кілька фаз: спочатку клітини не розмножуються, потім починають ділитися, причому швидкість розмноження весь час зростає; далі настає фаза, для якої характерна постійна швидкість ділення клітин; потім ця швидкість зменшується і настає відмирання клітин. Для отримання максимальної кількості клітин Б. вирощують в умовах т. Н. проточною культури; при цьому з посудини, в якому розмножуються Б., витікає певний обсяг культури; одночасно в судину додається в такій же кількості свіжа стерильна живильне середовище. При розмноженні Б. не в проточних, а в стаціонарних умовах відбувається зміна живильного середовища і накопичення в ній продуктів життєдіяльності Б., унаслідок чого змінюються і їх фізіологічні особливості. Так, молоді клітини Clostridium acetobutylicum не здатні утворювати ацетон; це властивість вони набувають в старішої культурі. Якщо спороносних Б. вирощувати в умови проточної культури, вони ділитимуться, але не даватимуть суперечку. При вирощуванні Б. на щільних поживних середовищах вони утворюють скупчення клітин різних розмірів, форми, кольору, звані колоніями.
Харчування. До складу клітин Б. входять ті ж біогенні елементи і мікроелементи, що і до складу клітин вищих рослин і тварин. Це С, N, О, Н, S, Р, К, Mg, Ca, Cl, Fe н ін. Крім білка, вуглеводів і жирів, Б. містять також РНК і велика кількість ДНК. Всі ці речовини можуть бути синтезовані тільки з речовин, що містяться в навколишньому середовищі. Як правило, через напівпроникну клітинну стінку і цитоплазматичну мембрану всередину Б. проходять тільки розчинні речовини. Під дією гідролітичних ферментів, що надходять з бактеріальних клітин назовні, відбувається розкладання складніших речовин (наприклад, крохмалю, целюлози) з утворенням розчинних продуктів (наприклад, моносахаров), засвоюваних Б. Як джерело азоту Б. можуть засвоювати білки, амінокислоти, амонійні солі , нітрати. Різні види Б. здатні утилізувати різні джерела азоту. Раніше вважали, що деякі патогенні (хвороботворні) і молочнокислі Б. можуть розвиватися лише в живильних середовищах з білками. Надалі з'ясувалося, що джерелом азоту для таких Б. можуть служити амонійні солі. Існує багато видів Б. з різних систематичних груп, які здатні засвоювати не тільки азот тих чи інших азотовмісних речовин, але і фіксувати азот атмосфери. До таких азотфиксирующим мікроорганізмам відносяться азотобактерии, мікобактерії, пурпурні фотосинтезирующие Б., а також бульбочкові бактерії (Див. Азотфіксация ). Джерелами мінерального живлення для Б. служать солі Р, S, Cl, К, Fe, Na, Ca; багато видів потребують також мікроелементів (Mo, Мn, Си, В, V і ін.). Для розмноження Б. необхідні також чинники зростання мікроорганізмів, до яких належать вітаміни групи В, біотин, пантотенова кислота, фолієва кислота та ін. Б., здатні синтезувати ці речовини, називаються ауксоавтотрофамі. До них відносяться псевдомоноси і багато інших неспороносні Б. При вирощуванні Б., не здатних до синтезу чинників зростання, їх додають в живильне середовище. Такі Б. називаються а у ксогетеротрофам і, до них відносяться, наприклад, різні молочнокислі Б. Джерелами вуглецю для Б. можуть служити вуглеводи, спирти, органічні кислоти, лігнін, хітин, вуглеводні, жири та ін. Здатність засвоювати вуглець з тих чи інших джерел у різних видів Б. варіює, і цим користуються для цілей систематики. Б., що засвоюють вуглець органічних сполук, називаються гетеротрофних. Б., що засвоюють вуглець вуглекислого газу атмосфери, називаються автотрофними. Види, що використовують для фіксації вуглекислоти енергію сонячних променів, наз. фотоавтотрофамі. Група Б., одержуючих енергію в результаті окислення таких неорганічних речовин, як аміак, нітрити, сірка, водень і ін., Здатних засвоювати вуглекислоту за рахунок енергії, що звільняється при окисленні вказаних неорганічних з'єднань, називаються хемоавтотрофов і, а сам процес асиміляції двоокису вуглецю, відкритий видатним російським мікробіологом С. Н. Виноградским, - хемосинтезом .
бактерії
Бакта е рії (грец. Bakterion - паличка), велика група (тип) мікроскопічних, переважно одноклітинних організмів, що володіють клітинною стінкою, що містять багато дезоксирибонуклеїнової кислоти (ДНК), що мають примітивне ядро, позбавлене видимих хромосом і оболонки, що не містять, як правило, хлорофілу і пластид, що розмножуються поперечним діленням (рідше перетяжкою або брунькуванням). Переважна кількість видів Б. має паличкоподібну форму. Однак до Б. відносять також мікроорганізми, що мають кулясту, ниткоподібну або звиту форму. Б. різноманітні по своїй фізіології, біохімічно дуже активні і поширені в грунті, воді, грунті водоймищ і пр. Б. не уявляють єдиної групи, а виникли різними шляхами. Деякі Б. (наприклад, нитчасті, азотобактер і ін.) Близькі до синезеленим водоростям , Ін. Б. споріднені променистим грибкам - актиноміцетам , Спірохети і деякі ін. Б. мають схожість з одноклітинними тваринами - найпростішими .
Б. беруть участь в круговороті речовин в природі, деякі з них викликають захворювання людини, тварин або рослин, застосовуються в різних галузях мікробіологічної промисловості. Науку, що вивчає Б., називають бактериологией . Це складова частина ширшої дисципліни - мікробіології , В завдання якої входить вивчення всіх сторін життєдіяльності не тільки Б., але і інших мікроорганізмів (дріжджі, цвілеві гриби, мікроскопічні водорості).
Людина використовував Б., ще не знаючи про їхнє існування. За допомогою заквасок, що містять Б., готували кисломолочні продукти, оцет, тісто і т.д. Вперше Б. побачив А. Левенгук - творець мікроскопа, досліджуючи рослинні настої і зубний наліт. До кінця 19 - початку 20 ст. було виділено велике число Б., що мешкають в грунті, воді, харчових продуктах і т.п., були відкриті багато видів хвороботворних Б. Класичні дослідження Л. Пастера в області фізіології Б. послужили основою для вивчення у них обміну речовин. Внесок у дослідження Б. внесли російські і радянські вчені С. Н. Виноградський , В. Л. Омелянський , Б. Л. Ісаченко , Які з'ясували роль Б. в круговороті речовин в природі, який робить можливим життя на Землі. Цей напрямок в мікробіології нерозривно пов'язане з розвитком геології, біогеохімії, ґрунтознавства, з вченням В. І. Вернадського про біосфері .
Морфологія і систематика Б. Розмір, форма, будова, рухливість Б. Діаметр кулястих Б. зазвичай дорівнює 1-2 мкм, ширина паличковидних форм коливається від 0,4 до 0,8 мкм, довжина дорівнює 2-5 мкм. Рідше зустрічаються дуже великі Б. Так, серобактерія Thiophysa macrophysa має 20 мкм в діаметрі, нитки інших серобактерий (Beggiatoa) видно неозброєним оком. Є також дуже дрібні Б., наприклад Bdellovibrio, що паразитують на Б. звичайних розмірів. Деякі Б., наприклад збудники плевропневмонії рогатої худоби, настільки дрібні, що невидимі в оптичний мікроскоп. Кулясті бактерії називаються кокками , Якщо ж вони розташовуються попарно, - диплококами (Рис. 1). Якщо коки розмножуються поперечним діленням і після ділення залишаються з'єднаними, утворюючи ланцюжки, то їх називають стрептококками (Рис. 2, 12). При діленні клітин в трьох взаємно перпендикулярних напрямках утворюються пакети кліток, типові для сарцин (Рис. 3). При розподілі коків в різних площинах виникають скупчення клітин у вигляді грона винограду, що характерно для стафілококів (Рис. 4). Паличкоподібні Б., що утворюють спори, називаються бацилами . Паличкоподібні форми можуть мати «обрубані» або опуклі кінці (рис. 5) і розташовуються окремо або, рідше, у вигляді ланцюжка (рис. 6). Б., що утворюють довгі нитки, - нитчасті бактерії , Мешкають переважно у воді. Б. у формі коми - вібріони (Рис. 7), покручені форми з грубими спіральними завитками - спірили (Рис. 8), з декількома рівномірними тонкими завитками - спірохети (Рис. 9).
Все Б. мають клітинну стінку (рис. 25, 26, 28). Вона виразно видно при приміщенні Б. у розчин кухонної солі; при цьому вміст клітини стискається і відстає від стінки - настає плазмоліз (Рис. 10). У ряду Б. стінка оточена слизової капсулою (рис. 27), присутність якої може бути встановлено при приміщенні таких Б. в розчин туші (рис. 11). При електронній мікроскопії видно, що клітинна стінка складається з декількох шарів (зазвичай трьох). До її складу входять мураміновая кислота, амінокислоти, ліпіди, глюкозамін і інші з'єднання. Хімічний склад клітинної стінки у різних систематичних груп, а також у Б., забарвлюються і не забарвлюються за Грамом, різний. Велику роль в обміні речовин грає цитоплазматична мембрана, яка перебуває під клітинною стінкою. У мембрані (рис. 25, 28) зосереджені численні ферментні системи бактеріальної клітини. У цитоплазмі є рибосоми , До складу яких входить РНК. Вміст нуклеїнових кислот у Б. коливається від 10 до 22% при різному ставленні РНК / ДНК (у кишкової палички воно дорівнює 2). За допомогою електронного мікроскопа встановлено присутність в клітці Б. ниток ДНК, що утворюють ядро, позбавлене оболонки, - т. Зв. нуклеоид (рис. 25, 26, 28). Будова ядра неоднаково у різних Б. Так, у «вищих», складніше організованих Б. (Myxobacteriales, Hyphomycrobiales), ядра легко можуть бути виявлені при мікроскопії забарвлених препаратів в оптичному мікроскопі (рис. 1). У багатьох Б. цитоплазма ущільнена на кінцях кліток і утворює забарвлюються полярні зерна (рис. 13; рис. 2). Клітини Б. містять запасні речовини: жирові включення (рис. 5), зерна глікогену, метахроматіна (рис. 4), гранульози (рис. 6), а також вакуолі, що містять рідину або газ. На відміну від грибів, Б. не містять мітохондрій , Що свідчить про більш примітивному будову Б. Багато Б. рухливі. Зазвичай такі форми мають довгі джгутики, що складаються з скорочувального білка (рис. 14). Завдяки хвилеподібним і спіральним рухам джгутиків клітка Б. переміщається. Види з одним джгутиком на полюсі клітини зв. монотріхі (рис. 15), клітини, що мають пучок джгутиків на кінці, - лофотрихи (рис. 16), Б., у яких джгутики розташовані по всій поверхні тіла, - перітріхі (рис. 17). У міксобактерій , Які також рухливі, джгутиків немає, і вони пересуваються в результаті набухання в довкіллі слизу, що виділяється клітинами (реактивний спосіб руху).
Життєвий цикл. Зміна морфології клітин Б. в часі дає уявлення про їх життєвому циклі. Так, багато аеробних і анаеробних Б. утворюють овальні або круглі блискучі спори . Такі види Б. називаються спороносними (або бацилами). Якщо суперечки великі і розташовуються в центрі клітини, то паличка набуває веретеноподібної форми (рис. 20); у інших видів спору розташовується на кінці палички, і тоді остання набуває форму булави (рис. 19) або барабанної палички. У багатьох спороносних бактерій діаметр спори невеликий, і тому при утворенні спори зберігається паличкоподібна форма Б. (рис. 18, 21). Надалі залишки вегетативної клітини руйнуються, і суперечки стає вільною (рис. 22). У кожній клітині утворюється тільки одна спору і, отже, спорообразование не можна розглядати як розмноження. Спори Б. дуже стійкі до дії високої температури і отруйних речовин. Потрапивши в сприятливе живильне середовище, спори проростають і з них виходять молоді паличкоподібні вегетативні клітини (рис. 23). Цикл розвитку Б. може бути різним. Так, мікобактерії розмножуються як діленням, так і брунькуванням (рис. 25). У міксобактерій вегетативні клітини стискаються, скорочуються і утворюють круглі або овальні мікроцисти (див. циста ), Які потім можуть проростати (рис. 24). Сполучені слизом мікроцисти утворюють тіла кулястої, грибовидной або Коралоподібних форми зеленого, рожевого або іншого кольору. В процесі зростання Б. можуть утворювати фільтрівні форми, що проходять через фільтри і дають в подальшому культури, подібні або тотожні з тими, в яких вони виникли.
Мінливість, фізіологічні і морфологічні властивості Б. можуть піддаватися змінам. Б. можуть втратити рухливість, здатність утворювати пігменти, давати суперечки, посилювати або зменшувати здатність до синтезу різних органічних сполук, змінювати форму і будову колоній на щільних поживних середовищах і т.д. Ці зміни можуть відбуватися мимовільно, т. Е. Безпрімененія відповідних зовнішніх впливів. Значно більше змінених форм виникає в результаті застосування мутагенів (ультрафіолетові промені, іонізуюча радіація, етиленімін або інших хімічних речовини). Кожна властивість Б. пов'язано з ДНК, т. Е. Контролюється відповідним геном . завдяки успіхам генетики мікроорганізмів встановлено місце розташування багатьох генів в нитки ДНК (див. Генетичні карти хромосом ). Виділивши ДНК з клітин-мутантів і додавши її в культуру іншого штаму, можна викликати (в результаті проникнення ДНК всередину клітин) спадкові зміни, називаються трансформацією . За допомогою мутагенів можуть бути отримані мутанти, цінні в практичному відношенні, в тому числі утворюють більшу кількість різних антибіотиків, амінокислот, вітамінів та інших біологічно активних речовин. За допомогою мутантів Б. були розшифровані шляху біосинтезу різних органічних сполук. Поступово змінюючи середовище проживання Б., можна адаптувати їх до нових умов існування. Так були отримані форми, стійкі до різних отрут, що розвиваються при незвичайній температурі або реакції середовища, і т.п .; так з'являються стійкі до деяких антибіотиків форми хвороботворних Б.
Систематика. Для з'ясування систематич. положення Б. визначають їх розміри, морфологію клітин, характер зростання чистої культури на різних питати. середовищах, форму, колір і характер поверхні колоній, що виростають на щільних середовищах. Встановлюють також характер розрідження Б. желатин (рис. 30), здатність їх згортати молоко, зброджувати різні вуглеводи, відновлювати нітрати, ооразовивать аміак, сірководень і індол при розкладанні білків і т.п. Надалі, маючи характеристику виділеної культури, визначають її систематичне положення. Б. підрозділяють на три класи. Перший клас - Eubacteria - об'єднує Б., що мають щільну клітинну стінку і не утворюють плодових тіл. В цьому класі розрізняють наступні порядки: 1) Eubacteriales - одноклітинні коки, неветвящиеся палички і спірально звиті форми; до цього порядку відносяться всі неспороносні і спороносні Б., фото-синтезують Б., спірохети та ін .; 2) Trichobacteriales - багатоклітинні нитчасті Б. з поперечними перегородками; 3) Ferribacteriales - одноклітинних нитчасті автотрофні железобактерии; 4) Thiobacteriales - одноклітинні автотрофні серобактерии. Другий клас - Myxobacteria - об'єднує Б. з тонкою клітинною стінкою і реактивним характером руху, що утворюють мікроцисти і плодові тіла різної форми. До третього класу - Hyphomicrobiales - відносять клітки, що дають довгі нитки, на кінцях яких утворюються нирки; відокремилися нирки рухливі. Сов. систематики виділяють на підставі еволюційних (філогенезі) даних великі групи, які об'єднують родинні форми. Так, наприклад, розгалужені мікобактерії не виділяються в самостійну групу, а об'єднані з актиноміцетами.
Фізіологія Б. Зростання, розмноження, розвиток. Після поділу Б. кожна з двох дочірніх бактеріальних клітин починає рости і досягає розмірів материнської. У цьому випадку говорять про зростання окремої клітини. Розмноження клітин, складових популяцію , Призводить до збільшення загального числа клітин. У цьому випадку говорять про зростання культури. При зростанні культури в рідкому поживному середовищі остання стає каламутною; чим більше клітин в культурі, тим вона мутніше. Про інтенсивність росту судять на підставі підрахунку клітин в 1 мл культури за допомогою мікроскопа або визначають за допомогою нефелометра ступінь каламутності живильного середовища. Визначаючи кількість клітин в різні періоди зростання культури, можна отримати криву зростання, що відображає кілька фаз: спочатку клітини не розмножуються, потім починають ділитися, причому швидкість розмноження весь час зростає; далі настає фаза, для якої характерна постійна швидкість ділення клітин; потім ця швидкість зменшується і настає відмирання клітин. Для отримання максимальної кількості клітин Б. вирощують в умовах т. Н. проточною культури; при цьому з посудини, в якому розмножуються Б., витікає певний обсяг культури; одночасно в судину додається в такій же кількості свіжа стерильна живильне середовище. При розмноженні Б. не в проточних, а в стаціонарних умовах відбувається зміна живильного середовища і накопичення в ній продуктів життєдіяльності Б., унаслідок чого змінюються і їх фізіологічні особливості. Так, молоді клітини Clostridium acetobutylicum не здатні утворювати ацетон; це властивість вони набувають в старішої культурі. Якщо спороносних Б. вирощувати в умови проточної культури, вони ділитимуться, але не даватимуть суперечку. При вирощуванні Б. на щільних поживних середовищах вони утворюють скупчення клітин різних розмірів, форми, кольору, звані колоніями.
Харчування. До складу клітин Б. входять ті ж біогенні елементи і мікроелементи, що і до складу клітин вищих рослин і тварин. Це С, N, О, Н, S, Р, К, Mg, Ca, Cl, Fe н ін. Крім білка, вуглеводів і жирів, Б. містять також РНК і велика кількість ДНК. Всі ці речовини можуть бути синтезовані тільки з речовин, що містяться в навколишньому середовищі. Як правило, через напівпроникну клітинну стінку і цитоплазматичну мембрану всередину Б. проходять тільки розчинні речовини. Під дією гідролітичних ферментів, що надходять з бактеріальних клітин назовні, відбувається розкладання складніших речовин (наприклад, крохмалю, целюлози) з утворенням розчинних продуктів (наприклад, моносахаров), засвоюваних Б. Як джерело азоту Б. можуть засвоювати білки, амінокислоти, амонійні солі , нітрати. Різні види Б. здатні утилізувати різні джерела азоту. Раніше вважали, що деякі патогенні (хвороботворні) і молочнокислі Б. можуть розвиватися лише в живильних середовищах з білками. Надалі з'ясувалося, що джерелом азоту для таких Б. можуть служити амонійні солі. Існує багато видів Б. з різних систематичних груп, які здатні засвоювати не тільки азот тих чи інших азотовмісних речовин, але і фіксувати азот атмосфери. До таких азотфиксирующим мікроорганізмам відносяться азотобактерии, мікобактерії, пурпурні фотосинтезирующие Б., а також бульбочкові бактерії (Див. Азотфіксация ). Джерелами мінерального живлення для Б. служать солі Р, S, Cl, К, Fe, Na, Ca; багато видів потребують також мікроелементів (Mo, Мn, Си, В, V і ін.). Для розмноження Б. необхідні також чинники зростання мікроорганізмів, до яких належать вітаміни групи В, біотин, пантотенова кислота, фолієва кислота та ін. Б., здатні синтезувати ці речовини, називаються ауксоавтотрофамі. До них відносяться псевдомоноси і багато інших неспороносні Б. При вирощуванні Б., не здатних до синтезу чинників зростання, їх додають в живильне середовище. Такі Б. називаються а у ксогетеротрофам і, до них відносяться, наприклад, різні молочнокислі Б. Джерелами вуглецю для Б. можуть служити вуглеводи, спирти, органічні кислоти, лігнін, хітин, вуглеводні, жири та ін. Здатність засвоювати вуглець з тих чи інших джерел у різних видів Б. варіює, і цим користуються для цілей систематики. Б., що засвоюють вуглець органічних сполук, називаються гетеротрофних. Б., що засвоюють вуглець вуглекислого газу атмосфери, називаються автотрофними. Види, що використовують для фіксації вуглекислоти енергію сонячних променів, наз. фотоавтотрофамі. Група Б., одержуючих енергію в результаті окислення таких неорганічних речовин, як аміак, нітрити, сірка, водень і ін., Здатних засвоювати вуглекислоту за рахунок енергії, що звільняється при окисленні вказаних неорганічних з'єднань, називаються хемоавтотрофов і, а сам процес асиміляції двоокису вуглецю, відкритий видатним російським мікробіологом С. Н. Виноградским, - хемосинтезом .
бактерії
Бакта е рії (грец. Bakterion - паличка), велика група (тип) мікроскопічних, переважно одноклітинних організмів, що володіють клітинною стінкою, що містять багато дезоксирибонуклеїнової кислоти (ДНК), що мають примітивне ядро, позбавлене видимих хромосом і оболонки, що не містять, як правило, хлорофілу і пластид, що розмножуються поперечним діленням (рідше перетяжкою або брунькуванням). Переважна кількість видів Б. має паличкоподібну форму. Однак до Б. відносять також мікроорганізми, що мають кулясту, ниткоподібну або звиту форму. Б. різноманітні по своїй фізіології, біохімічно дуже активні і поширені в грунті, воді, грунті водоймищ і пр. Б. не уявляють єдиної групи, а виникли різними шляхами. Деякі Б. (наприклад, нитчасті, азотобактер і ін.) Близькі до синезеленим водоростям , Ін. Б. споріднені променистим грибкам - актиноміцетам , Спірохети і деякі ін. Б. мають схожість з одноклітинними тваринами - найпростішими .
Б. беруть участь в круговороті речовин в природі, деякі з них викликають захворювання людини, тварин або рослин, застосовуються в різних галузях мікробіологічної промисловості. Науку, що вивчає Б., називають бактериологией . Це складова частина ширшої дисципліни - мікробіології , В завдання якої входить вивчення всіх сторін життєдіяльності не тільки Б., але і інших мікроорганізмів (дріжджі, цвілеві гриби, мікроскопічні водорості).
Людина використовував Б., ще не знаючи про їхнє існування. За допомогою заквасок, що містять Б., готували кисломолочні продукти, оцет, тісто і т.д. Вперше Б. побачив А. Левенгук - творець мікроскопа, досліджуючи рослинні настої і зубний наліт. До кінця 19 - початку 20 ст. було виділено велике число Б., що мешкають в грунті, воді, харчових продуктах і т.п., були відкриті багато видів хвороботворних Б. Класичні дослідження Л. Пастера в області фізіології Б. послужили основою для вивчення у них обміну речовин. Внесок у дослідження Б. внесли російські і радянські вчені С. Н. Виноградський , В. Л. Омелянський , Б. Л. Ісаченко , Які з'ясували роль Б. в круговороті речовин в природі, який робить можливим життя на Землі. Цей напрямок в мікробіології нерозривно пов'язане з розвитком геології, біогеохімії, ґрунтознавства, з вченням В. І. Вернадського про біосфері .
Морфологія і систематика Б. Розмір, форма, будова, рухливість Б. Діаметр кулястих Б. зазвичай дорівнює 1-2 мкм, ширина паличковидних форм коливається від 0,4 до 0,8 мкм, довжина дорівнює 2-5 мкм. Рідше зустрічаються дуже великі Б. Так, серобактерія Thiophysa macrophysa має 20 мкм в діаметрі, нитки інших серобактерий (Beggiatoa) видно неозброєним оком. Є також дуже дрібні Б., наприклад Bdellovibrio, що паразитують на Б. звичайних розмірів. Деякі Б., наприклад збудники плевропневмонії рогатої худоби, настільки дрібні, що невидимі в оптичний мікроскоп. Кулясті бактерії називаються кокками , Якщо ж вони розташовуються попарно, - диплококами (Рис. 1). Якщо коки розмножуються поперечним діленням і після ділення залишаються з'єднаними, утворюючи ланцюжки, то їх називають стрептококками (Рис. 2, 12). При діленні клітин в трьох взаємно перпендикулярних напрямках утворюються пакети кліток, типові для сарцин (Рис. 3). При розподілі коків в різних площинах виникають скупчення клітин у вигляді грона винограду, що характерно для стафілококів (Рис. 4). Паличкоподібні Б., що утворюють спори, називаються бацилами . Паличкоподібні форми можуть мати «обрубані» або опуклі кінці (рис. 5) і розташовуються окремо або, рідше, у вигляді ланцюжка (рис. 6). Б., що утворюють довгі нитки, - нитчасті бактерії , Мешкають переважно у воді. Б. у формі коми - вібріони (Рис. 7), покручені форми з грубими спіральними завитками - спірили (Рис. 8), з декількома рівномірними тонкими завитками - спірохети (Рис. 9).
Все Б. мають клітинну стінку (рис. 25, 26, 28). Вона виразно видно при приміщенні Б. у розчин кухонної солі; при цьому вміст клітини стискається і відстає від стінки - настає плазмоліз (Рис. 10). У ряду Б. стінка оточена слизової капсулою (рис. 27), присутність якої може бути встановлено при приміщенні таких Б. в розчин туші (рис. 11). При електронній мікроскопії видно, що клітинна стінка складається з декількох шарів (зазвичай трьох). До її складу входять мураміновая кислота, амінокислоти, ліпіди, глюкозамін і інші з'єднання. Хімічний склад клітинної стінки у різних систематичних груп, а також у Б., забарвлюються і не забарвлюються за Грамом, різний. Велику роль в обміні речовин грає цитоплазматична мембрана, яка перебуває під клітинною стінкою. У мембрані (рис. 25, 28) зосереджені численні ферментні системи бактеріальної клітини. У цитоплазмі є рибосоми , До складу яких входить РНК. Вміст нуклеїнових кислот у Б. коливається від 10 до 22% при різному ставленні РНК / ДНК (у кишкової палички воно дорівнює 2). За допомогою електронного мікроскопа встановлено присутність в клітці Б. ниток ДНК, що утворюють ядро, позбавлене оболонки, - т. Зв. нуклеоид (рис. 25, 26, 28). Будова ядра неоднаково у різних Б. Так, у «вищих», складніше організованих Б. (Myxobacteriales, Hyphomycrobiales), ядра легко можуть бути виявлені при мікроскопії забарвлених препаратів в оптичному мікроскопі (рис. 1). У багатьох Б. цитоплазма ущільнена на кінцях кліток і утворює забарвлюються полярні зерна (рис. 13; рис. 2). Клітини Б. містять запасні речовини: жирові включення (рис. 5), зерна глікогену, метахроматіна (рис. 4), гранульози (рис. 6), а також вакуолі, що містять рідину або газ. На відміну від грибів, Б. не містять мітохондрій , Що свідчить про більш примітивному будову Б. Багато Б. рухливі. Зазвичай такі форми мають довгі джгутики, що складаються з скорочувального білка (рис. 14). Завдяки хвилеподібним і спіральним рухам джгутиків клітка Б. переміщається. Види з одним джгутиком на полюсі клітини зв. монотріхі (рис. 15), клітини, що мають пучок джгутиків на кінці, - лофотрихи (рис. 16), Б., у яких джгутики розташовані по всій поверхні тіла, - перітріхі (рис. 17). У міксобактерій , Які також рухливі, джгутиків немає, і вони пересуваються в результаті набухання в довкіллі слизу, що виділяється клітинами (реактивний спосіб руху).
Життєвий цикл. Зміна морфології клітин Б. в часі дає уявлення про їх життєвому циклі. Так, багато аеробних і анаеробних Б. утворюють овальні або круглі блискучі спори . Такі види Б. називаються спороносними (або бацилами). Якщо суперечки великі і розташовуються в центрі клітини, то паличка набуває веретеноподібної форми (рис. 20); у інших видів спору розташовується на кінці палички, і тоді остання набуває форму булави (рис. 19) або барабанної палички. У багатьох спороносних бактерій діаметр спори невеликий, і тому при утворенні спори зберігається паличкоподібна форма Б. (рис. 18, 21). Надалі залишки вегетативної клітини руйнуються, і суперечки стає вільною (рис. 22). У кожній клітині утворюється тільки одна спору і, отже, спорообразование не можна розглядати як розмноження. Спори Б. дуже стійкі до дії високої температури і отруйних речовин. Потрапивши в сприятливе живильне середовище, спори проростають і з них виходять молоді паличкоподібні вегетативні клітини (рис. 23). Цикл розвитку Б. може бути різним. Так, мікобактерії розмножуються як діленням, так і брунькуванням (рис. 25). У міксобактерій вегетативні клітини стискаються, скорочуються і утворюють круглі або овальні мікроцисти (див. циста ), Які потім можуть проростати (рис. 24). Сполучені слизом мікроцисти утворюють тіла кулястої, грибовидной або Коралоподібних форми зеленого, рожевого або іншого кольору. В процесі зростання Б. можуть утворювати фільтрівні форми, що проходять через фільтри і дають в подальшому культури, подібні або тотожні з тими, в яких вони виникли.
Мінливість, фізіологічні і морфологічні властивості Б. можуть піддаватися змінам. Б. можуть втратити рухливість, здатність утворювати пігменти, давати суперечки, посилювати або зменшувати здатність до синтезу різних органічних сполук, змінювати форму і будову колоній на щільних поживних середовищах і т.д. Ці зміни можуть відбуватися мимовільно, т. Е. Безпрімененія відповідних зовнішніх впливів. Значно більше змінених форм виникає в результаті застосування мутагенів (ультрафіолетові промені, іонізуюча радіація, етиленімін або інших хімічних речовини). Кожна властивість Б. пов'язано з ДНК, т. Е. Контролюється відповідним геном . завдяки успіхам генетики мікроорганізмів встановлено місце розташування багатьох генів в нитки ДНК (див. Генетичні карти хромосом ). Виділивши ДНК з клітин-мутантів і додавши її в культуру іншого штаму, можна викликати (в результаті проникнення ДНК всередину клітин) спадкові зміни, називаються трансформацією . За допомогою мутагенів можуть бути отримані мутанти, цінні в практичному відношенні, в тому числі утворюють більшу кількість різних антибіотиків, амінокислот, вітамінів та інших біологічно активних речовин. За допомогою мутантів Б. були розшифровані шляху біосинтезу різних органічних сполук. Поступово змінюючи середовище проживання Б., можна адаптувати їх до нових умов існування. Так були отримані форми, стійкі до різних отрут, що розвиваються при незвичайній температурі або реакції середовища, і т.п .; так з'являються стійкі до деяких антибіотиків форми хвороботворних Б.
Систематика. Для з'ясування систематич. положення Б. визначають їх розміри, морфологію клітин, характер зростання чистої культури на різних питати. середовищах, форму, колір і характер поверхні колоній, що виростають на щільних середовищах. Встановлюють також характер розрідження Б. желатин (рис. 30), здатність їх згортати молоко, зброджувати різні вуглеводи, відновлювати нітрати, ооразовивать аміак, сірководень і індол при розкладанні білків і т.п. Надалі, маючи характеристику виділеної культури, визначають її систематичне положення. Б. підрозділяють на три класи. Перший клас - Eubacteria - об'єднує Б., що мають щільну клітинну стінку і не утворюють плодових тіл. В цьому класі розрізняють наступні порядки: 1) Eubacteriales - одноклітинні коки, неветвящиеся палички і спірально звиті форми; до цього порядку відносяться всі неспороносні і спороносні Б., фото-синтезують Б., спірохети та ін .; 2) Trichobacteriales - багатоклітинні нитчасті Б. з поперечними перегородками; 3) Ferribacteriales - одноклітинних нитчасті автотрофні железобактерии; 4) Thiobacteriales - одноклітинні автотрофні серобактерии. Другий клас - Myxobacteria - об'єднує Б. з тонкою клітинною стінкою і реактивним характером руху, що утворюють мікроцисти і плодові тіла різної форми. До третього класу - Hyphomicrobiales - відносять клітки, що дають довгі нитки, на кінцях яких утворюються нирки; відокремилися нирки рухливі. Сов. систематики виділяють на підставі еволюційних (філогенезі) даних великі групи, які об'єднують родинні форми. Так, наприклад, розгалужені мікобактерії не виділяються в самостійну групу, а об'єднані з актиноміцетами.
Фізіологія Б. Зростання, розмноження, розвиток. Після поділу Б. кожна з двох дочірніх бактеріальних клітин починає рости і досягає розмірів материнської. У цьому випадку говорять про зростання окремої клітини. Розмноження клітин, складових популяцію , Призводить до збільшення загального числа клітин. У цьому випадку говорять про зростання культури. При зростанні культури в рідкому поживному середовищі остання стає каламутною; чим більше клітин в культурі, тим вона мутніше. Про інтенсивність росту судять на підставі підрахунку клітин в 1 мл культури за допомогою мікроскопа або визначають за допомогою нефелометра ступінь каламутності живильного середовища. Визначаючи кількість клітин в різні періоди зростання культури, можна отримати криву зростання, що відображає кілька фаз: спочатку клітини не розмножуються, потім починають ділитися, причому швидкість розмноження весь час зростає; далі настає фаза, для якої характерна постійна швидкість ділення клітин; потім ця швидкість зменшується і настає відмирання клітин. Для отримання максимальної кількості клітин Б. вирощують в умовах т. Н. проточною культури; при цьому з посудини, в якому розмножуються Б., витікає певний обсяг культури; одночасно в судину додається в такій же кількості свіжа стерильна живильне середовище. При розмноженні Б. не в проточних, а в стаціонарних умовах відбувається зміна живильного середовища і накопичення в ній продуктів життєдіяльності Б., унаслідок чого змінюються і їх фізіологічні особливості. Так, молоді клітини Clostridium acetobutylicum не здатні утворювати ацетон; це властивість вони набувають в старішої культурі. Якщо спороносних Б. вирощувати в умови проточної культури, вони ділитимуться, але не даватимуть суперечку. При вирощуванні Б. на щільних поживних середовищах вони утворюють скупчення клітин різних розмірів, форми, кольору, звані колоніями.
Харчування. До складу клітин Б. входять ті ж біогенні елементи і мікроелементи, що і до складу клітин вищих рослин і тварин. Це С, N, О, Н, S, Р, К, Mg, Ca, Cl, Fe н ін. Крім білка, вуглеводів і жирів, Б. містять також РНК і велика кількість ДНК. Всі ці речовини можуть бути синтезовані тільки з речовин, що містяться в навколишньому середовищі. Як правило, через напівпроникну клітинну стінку і цитоплазматичну мембрану всередину Б. проходять тільки розчинні речовини. Під дією гідролітичних ферментів, що надходять з бактеріальних клітин назовні, відбувається розкладання складніших речовин (наприклад, крохмалю, целюлози) з утворенням розчинних продуктів (наприклад, моносахаров), засвоюваних Б. Як джерело азоту Б. можуть засвоювати білки, амінокислоти, амонійні солі , нітрати. Різні види Б. здатні утилізувати різні джерела азоту. Раніше вважали, що деякі патогенні (хвороботворні) і молочнокислі Б. можуть розвиватися лише в живильних середовищах з білками. Надалі з'ясувалося, що джерелом азоту для таких Б. можуть служити амонійні солі. Існує багато видів Б. з різних систематичних груп, які здатні засвоювати не тільки азот тих чи інших азотовмісних речовин, але і фіксувати азот атмосфери. До таких азотфиксирующим мікроорганізмам відносяться азотобактерии, мікобактерії, пурпурні фотосинтезирующие Б., а також бульбочкові бактерії (Див. Азотфіксация ). Джерелами мінерального живлення для Б. служать солі Р, S, Cl, К, Fe, Na, Ca; багато видів потребують також мікроелементів (Mo, Мn, Си, В, V і ін.). Для розмноження Б. необхідні також чинники зростання мікроорганізмів, до яких належать вітаміни групи В, біотин, пантотенова кислота, фолієва кислота та ін. Б., здатні синтезувати ці речовини, називаються ауксоавтотрофамі. До них відносяться псевдомоноси і багато інших неспороносні Б. При вирощуванні Б., не здатних до синтезу чинників зростання, їх додають в живильне середовище. Такі Б. називаються а у ксогетеротрофам і, до них відносяться, наприклад, різні молочнокислі Б. Джерелами вуглецю для Б. можуть служити вуглеводи, спирти, органічні кислоти, лігнін, хітин, вуглеводні, жири та ін. Здатність засвоювати вуглець з тих чи інших джерел у різних видів Б. варіює, і цим користуються для цілей систематики. Б., що засвоюють вуглець органічних сполук, називаються гетеротрофних. Б., що засвоюють вуглець вуглекислого газу атмосфери, називаються автотрофними. Види, що використовують для фіксації вуглекислоти енергію сонячних променів, наз. фотоавтотрофамі. Група Б., одержуючих енергію в результаті окислення таких неорганічних речовин, як аміак, нітрити, сірка, водень і ін., Здатних засвоювати вуглекислоту за рахунок енергії, що звільняється при окисленні вказаних неорганічних з'єднань, називаються хемоавтотрофов і, а сам процес асиміляції двоокису вуглецю, відкритий видатним російським мікробіологом С. Н. Виноградским, - хемосинтезом .
бактерії
Бакта е рії (грец. Bakterion - паличка), велика група (тип) мікроскопічних, переважно одноклітинних організмів, що володіють клітинною стінкою, що містять багато дезоксирибонуклеїнової кислоти (ДНК), що мають примітивне ядро, позбавлене видимих хромосом і оболонки, що не містять, як правило, хлорофілу і пластид, що розмножуються поперечним діленням (рідше перетяжкою або брунькуванням). Переважна кількість видів Б. має паличкоподібну форму. Однак до Б. відносять також мікроорганізми, що мають кулясту, ниткоподібну або звиту форму. Б. різноманітні по своїй фізіології, біохімічно дуже активні і поширені в грунті, воді, грунті водоймищ і пр. Б. не уявляють єдиної групи, а виникли різними шляхами. Деякі Б. (наприклад, нитчасті, азотобактер і ін.) Близькі до синезеленим водоростям , Ін. Б. споріднені променистим грибкам - актиноміцетам , Спірохети і деякі ін. Б. мають схожість з одноклітинними тваринами - найпростішими .
Б. беруть участь в круговороті речовин в природі, деякі з них викликають захворювання людини, тварин або рослин, застосовуються в різних галузях мікробіологічної промисловості. Науку, що вивчає Б., називають бактериологией . Це складова частина ширшої дисципліни - мікробіології , В завдання якої входить вивчення всіх сторін життєдіяльності не тільки Б., але і інших мікроорганізмів (дріжджі, цвілеві гриби, мікроскопічні водорості).
Людина використовував Б., ще не знаючи про їхнє існування. За допомогою заквасок, що містять Б., готували кисломолочні продукти, оцет, тісто і т.д. Вперше Б. побачив А. Левенгук - творець мікроскопа, досліджуючи рослинні настої і зубний наліт. До кінця 19 - початку 20 ст. було виділено велике число Б., що мешкають в грунті, воді, харчових продуктах і т.п., були відкриті багато видів хвороботворних Б. Класичні дослідження Л. Пастера в області фізіології Б. послужили основою для вивчення у них обміну речовин. Внесок у дослідження Б. внесли російські і радянські вчені С. Н. Виноградський , В. Л. Омелянський , Б. Л. Ісаченко , Які з'ясували роль Б. в круговороті речовин в природі, який робить можливим життя на Землі. Цей напрямок в мікробіології нерозривно пов'язане з розвитком геології, біогеохімії, ґрунтознавства, з вченням В. І. Вернадського про біосфері .
Морфологія і систематика Б. Розмір, форма, будова, рухливість Б. Діаметр кулястих Б. зазвичай дорівнює 1-2 мкм, ширина паличковидних форм коливається від 0,4 до 0,8 мкм, довжина дорівнює 2-5 мкм. Рідше зустрічаються дуже великі Б. Так, серобактерія Thiophysa macrophysa має 20 мкм в діаметрі, нитки інших серобактерий (Beggiatoa) видно неозброєним оком. Є також дуже дрібні Б., наприклад Bdellovibrio, що паразитують на Б. звичайних розмірів. Деякі Б., наприклад збудники плевропневмонії рогатої худоби, настільки дрібні, що невидимі в оптичний мікроскоп. Кулясті бактерії називаються кокками , Якщо ж вони розташовуються попарно, - диплококами (Рис. 1). Якщо коки розмножуються поперечним діленням і після ділення залишаються з'єднаними, утворюючи ланцюжки, то їх називають стрептококками (Рис. 2, 12). При діленні клітин в трьох взаємно перпендикулярних напрямках утворюються пакети кліток, типові для сарцин (Рис. 3). При розподілі коків в різних площинах виникають скупчення клітин у вигляді грона винограду, що характерно для стафілококів (Рис. 4). Паличкоподібні Б., що утворюють спори, називаються бацилами . Паличкоподібні форми можуть мати «обрубані» або опуклі кінці (рис. 5) і розташовуються окремо або, рідше, у вигляді ланцюжка (рис. 6). Б., що утворюють довгі нитки, - нитчасті бактерії , Мешкають переважно у воді. Б. у формі коми - вібріони (Рис. 7), покручені форми з грубими спіральними завитками - спірили (Рис. 8), з декількома рівномірними тонкими завитками - спірохети (Рис. 9).
Все Б. мають клітинну стінку (рис. 25, 26, 28). Вона виразно видно при приміщенні Б. у розчин кухонної солі; при цьому вміст клітини стискається і відстає від стінки - настає плазмоліз (Рис. 10). У ряду Б. стінка оточена слизової капсулою (рис. 27), присутність якої може бути встановлено при приміщенні таких Б. в розчин туші (рис. 11). При електронній мікроскопії видно, що клітинна стінка складається з декількох шарів (зазвичай трьох). До її складу входять мураміновая кислота, амінокислоти, ліпіди, глюкозамін і інші з'єднання. Хімічний склад клітинної стінки у різних систематичних груп, а також у Б., забарвлюються і не забарвлюються за Грамом, різний. Велику роль в обміні речовин грає цитоплазматична мембрана, яка перебуває під клітинною стінкою. У мембрані (рис. 25, 28) зосереджені численні ферментні системи бактеріальної клітини. У цитоплазмі є рибосоми , До складу яких входить РНК. Вміст нуклеїнових кислот у Б. коливається від 10 до 22% при різному ставленні РНК / ДНК (у кишкової палички воно дорівнює 2). За допомогою електронного мікроскопа встановлено присутність в клітці Б. ниток ДНК, що утворюють ядро, позбавлене оболонки, - т. Зв. нуклеоид (рис. 25, 26, 28). Будова ядра неоднаково у різних Б. Так, у «вищих», складніше організованих Б. (Myxobacteriales, Hyphomycrobiales), ядра легко можуть бути виявлені при мікроскопії забарвлених препаратів в оптичному мікроскопі (рис. 1). У багатьох Б. цитоплазма ущільнена на кінцях кліток і утворює забарвлюються полярні зерна (рис. 13; рис. 2). Клітини Б. містять запасні речовини: жирові включення (рис. 5), зерна глікогену, метахроматіна (рис. 4), гранульози (рис. 6), а також вакуолі, що містять рідину або газ. На відміну від грибів, Б. не містять мітохондрій , Що свідчить про більш примітивному будову Б. Багато Б. рухливі. Зазвичай такі форми мають довгі джгутики, що складаються з скорочувального білка (рис. 14). Завдяки хвилеподібним і спіральним рухам джгутиків клітка Б. переміщається. Види з одним джгутиком на полюсі клітини зв. монотріхі (рис. 15), клітини, що мають пучок джгутиків на кінці, - лофотрихи (рис. 16), Б., у яких джгутики розташовані по всій поверхні тіла, - перітріхі (рис. 17). У міксобактерій , Які також рухливі, джгутиків немає, і вони пересуваються в результаті набухання в довкіллі слизу, що виділяється клітинами (реактивний спосіб руху).
Життєвий цикл. Зміна морфології клітин Б. в часі дає уявлення про їх життєвому циклі. Так, багато аеробних і анаеробних Б. утворюють овальні або круглі блискучі спори . Такі види Б. називаються спороносними (або бацилами). Якщо суперечки великі і розташовуються в центрі клітини, то паличка набуває веретеноподібної форми (рис. 20); у інших видів спору розташовується на кінці палички, і тоді остання набуває форму булави (рис. 19) або барабанної палички. У багатьох спороносних бактерій діаметр спори невеликий, і тому при утворенні спори зберігається паличкоподібна форма Б. (рис. 18, 21). Надалі залишки вегетативної клітини руйнуються, і суперечки стає вільною (рис. 22). У кожній клітині утворюється тільки одна спору і, отже, спорообразование не можна розглядати як розмноження. Спори Б. дуже стійкі до дії високої температури і отруйних речовин. Потрапивши в сприятливе живильне середовище, спори проростають і з них виходять молоді паличкоподібні вегетативні клітини (рис. 23). Цикл розвитку Б. може бути різним. Так, мікобактерії розмножуються як діленням, так і брунькуванням (рис. 25). У міксобактерій вегетативні клітини стискаються, скорочуються і утворюють круглі або овальні мікроцисти (див. циста ), Які потім можуть проростати (рис. 24). Сполучені слизом мікроцисти утворюють тіла кулястої, грибовидной або Коралоподібних форми зеленого, рожевого або іншого кольору. В процесі зростання Б. можуть утворювати фільтрівні форми, що проходять через фільтри і дають в подальшому культури, подібні або тотожні з тими, в яких вони виникли.
Мінливість, фізіологічні і морфологічні властивості Б. можуть піддаватися змінам. Б. можуть втратити рухливість, здатність утворювати пігменти, давати суперечки, посилювати або зменшувати здатність до синтезу різних органічних сполук, змінювати форму і будову колоній на щільних поживних середовищах і т.д. Ці зміни можуть відбуватися мимовільно, т. Е. Безпрімененія відповідних зовнішніх впливів. Значно більше змінених форм виникає в результаті застосування мутагенів (ультрафіолетові промені, іонізуюча радіація, етиленімін або інших хімічних речовини). Кожна властивість Б. пов'язано з ДНК, т. Е. Контролюється відповідним геном . завдяки успіхам генетики мікроорганізмів встановлено місце розташування багатьох генів в нитки ДНК (див. Генетичні карти хромосом ). Виділивши ДНК з клітин-мутантів і додавши її в культуру іншого штаму, можна викликати (в результаті проникнення ДНК всередину клітин) спадкові зміни, називаються трансформацією . За допомогою мутагенів можуть бути отримані мутанти, цінні в практичному відношенні, в тому числі утворюють більшу кількість різних антибіотиків, амінокислот, вітамінів та інших біологічно активних речовин. За допомогою мутантів Б. були розшифровані шляху біосинтезу різних органічних сполук. Поступово змінюючи середовище проживання Б., можна адаптувати їх до нових умов існування. Так були отримані форми, стійкі до різних отрут, що розвиваються при незвичайній температурі або реакції середовища, і т.п .; так з'являються стійкі до деяких антибіотиків форми хвороботворних Б.
Систематика. Для з'ясування систематич. положення Б. визначають їх розміри, морфологію клітин, характер зростання чистої культури на різних питати. середовищах, форму, колір і характер поверхні колоній, що виростають на щільних середовищах. Встановлюють також характер розрідження Б. желатин (рис. 30), здатність їх згортати молоко, зброджувати різні вуглеводи, відновлювати нітрати, ооразовивать аміак, сірководень і індол при розкладанні білків і т.п. Надалі, маючи характеристику виділеної культури, визначають її систематичне положення. Б. підрозділяють на три класи. Перший клас - Eubacteria - об'єднує Б., що мають щільну клітинну стінку і не утворюють плодових тіл. В цьому класі розрізняють наступні порядки: 1) Eubacteriales - одноклітинні коки, неветвящиеся палички і спірально звиті форми; до цього порядку відносяться всі неспороносні і спороносні Б., фото-синтезують Б., спірохети та ін .; 2) Trichobacteriales - багатоклітинні нитчасті Б. з поперечними перегородками; 3) Ferribacteriales - одноклітинних нитчасті автотрофні железобактерии; 4) Thiobacteriales - одноклітинні автотрофні серобактерии. Другий клас - Myxobacteria - об'єднує Б. з тонкою клітинною стінкою і реактивним характером руху, що утворюють мікроцисти і плодові тіла різної форми. До третього класу - Hyphomicrobiales - відносять клітки, що дають довгі нитки, на кінцях яких утворюються нирки; відокремилися нирки рухливі. Сов. систематики виділяють на підставі еволюційних (філогенезі) даних великі групи, які об'єднують родинні форми. Так, наприклад, розгалужені мікобактерії не виділяються в самостійну групу, а об'єднані з актиноміцетами.
Фізіологія Б. Зростання, розмноження, розвиток. Після поділу Б. кожна з двох дочірніх бактеріальних клітин починає рости і досягає розмірів материнської. У цьому випадку говорять про зростання окремої клітини. Розмноження клітин, складових популяцію , Призводить до збільшення загального числа клітин. У цьому випадку говорять про зростання культури. При зростанні культури в рідкому поживному середовищі остання стає каламутною; чим більше клітин в культурі, тим вона мутніше. Про інтенсивність росту судять на підставі підрахунку клітин в 1 мл культури за допомогою мікроскопа або визначають за допомогою нефелометра ступінь каламутності живильного середовища. Визначаючи кількість клітин в різні періоди зростання культури, можна отримати криву зростання, що відображає кілька фаз: спочатку клітини не розмножуються, потім починають ділитися, причому швидкість розмноження весь час зростає; далі настає фаза, для якої характерна постійна швидкість ділення клітин; потім ця швидкість зменшується і настає відмирання клітин. Для отримання максимальної кількості клітин Б. вирощують в умовах т. Н. проточною культури; при цьому з посудини, в якому розмножуються Б., витікає певний обсяг культури; одночасно в судину додається в такій же кількості свіжа стерильна живильне середовище. При розмноженні Б. не в проточних, а в стаціонарних умовах відбувається зміна живильного середовища і накопичення в ній продуктів життєдіяльності Б., унаслідок чого змінюються і їх фізіологічні особливості. Так, молоді клітини Clostridium acetobutylicum не здатні утворювати ацетон; це властивість вони набувають в старішої культурі. Якщо спороносних Б. вирощувати в умови проточної культури, вони ділитимуться, але не даватимуть суперечку. При вирощуванні Б. на щільних поживних середовищах вони утворюють скупчення клітин різних розмірів, форми, кольору, звані колоніями.
Харчування. До складу клітин Б. входять ті ж біогенні елементи і мікроелементи, що і до складу клітин вищих рослин і тварин. Це С, N, О, Н, S, Р, К, Mg, Ca, Cl, Fe н ін. Крім білка, вуглеводів і жирів, Б. містять також РНК і велика кількість ДНК. Всі ці речовини можуть бути синтезовані тільки з речовин, що містяться в навколишньому середовищі. Як правило, через напівпроникну клітинну стінку і цитоплазматичну мембрану всередину Б. проходять тільки розчинні речовини. Під дією гідролітичних ферментів, що надходять з бактеріальних клітин назовні, відбувається розкладання складніших речовин (наприклад, крохмалю, целюлози) з утворенням розчинних продуктів (наприклад, моносахаров), засвоюваних Б. Як джерело азоту Б. можуть засвоювати білки, амінокислоти, амонійні солі , нітрати. Різні види Б. здатні утилізувати різні джерела азоту. Раніше вважали, що деякі патогенні (хвороботворні) і молочнокислі Б. можуть розвиватися лише в живильних середовищах з білками. Надалі з'ясувалося, що джерелом азоту для таких Б. можуть служити амонійні солі. Існує багато видів Б. з різних систематичних груп, які здатні засвоювати не тільки азот тих чи інших азотовмісних речовин, але і фіксувати азот атмосфери. До таких азотфиксирующим мікроорганізмам відносяться азотобактерии, мікобактерії, пурпурні фотосинтезирующие Б., а також бульбочкові бактерії (Див. Азотфіксация ). Джерелами мінерального живлення для Б. служать солі Р, S, Cl, К, Fe, Na, Ca; багато видів потребують також мікроелементів (Mo, Мn, Си, В, V і ін.). Для розмноження Б. необхідні також чинники зростання мікроорганізмів, до яких належать вітаміни групи В, біотин, пантотенова кислота, фолієва кислота та ін. Б., здатні синтезувати ці речовини, називаються ауксоавтотрофамі. До них відносяться псевдомоноси і багато інших неспороносні Б. При вирощуванні Б., не здатних до синтезу чинників зростання, їх додають в живильне середовище. Такі Б. називаються а у ксогетеротрофам і, до них відносяться, наприклад, різні молочнокислі Б. Джерелами вуглецю для Б. можуть служити вуглеводи, спирти, органічні кислоти, лігнін, хітин, вуглеводні, жири та ін. Здатність засвоювати вуглець з тих чи інших джерел у різних видів Б. варіює, і цим користуються для цілей систематики. Б., що засвоюють вуглець органічних сполук, називаються гетеротрофних. Б., що засвоюють вуглець вуглекислого газу атмосфери, називаються автотрофними. Види, що використовують для фіксації вуглекислоти енергію сонячних променів, наз. фотоавтотрофамі. Група Б., одержуючих енергію в результаті окислення таких неорганічних речовин, як аміак, нітрити, сірка, водень і ін., Здатних засвоювати вуглекислоту за рахунок енергії, що звільняється при окисленні вказаних неорганічних з'єднань, називаються хемоавтотрофов і, а сам процес асиміляції двоокису вуглецю, відкритий видатним російським мікробіологом С. Н. Виноградским, - хемосинтезом .
бактерії
Бакта е рії (грец. Bakterion - паличка), велика група (тип) мікроскопічних, переважно одноклітинних організмів, що володіють клітинною стінкою, що містять багато дезоксирибонуклеїнової кислоти (ДНК), що мають примітивне ядро, позбавлене видимих хромосом і оболонки, що не містять, як правило, хлорофілу і пластид, що розмножуються поперечним діленням (рідше перетяжкою або брунькуванням). Переважна кількість видів Б. має паличкоподібну форму. Однак до Б. відносять також мікроорганізми, що мають кулясту, ниткоподібну або звиту форму. Б. різноманітні по своїй фізіології, біохімічно дуже активні і поширені в грунті, воді, грунті водоймищ і пр. Б. не уявляють єдиної групи, а виникли різними шляхами. Деякі Б. (наприклад, нитчасті, азотобактер і ін.) Близькі до синезеленим водоростям , Ін. Б. споріднені променистим грибкам - актиноміцетам , Спірохети і деякі ін. Б. мають схожість з одноклітинними тваринами - найпростішими .
Б. беруть участь в круговороті речовин в природі, деякі з них викликають захворювання людини, тварин або рослин, застосовуються в різних галузях мікробіологічної промисловості. Науку, що вивчає Б., називають бактериологией . Це складова частина ширшої дисципліни - мікробіології , В завдання якої входить вивчення всіх сторін життєдіяльності не тільки Б., але і інших мікроорганізмів (дріжджі, цвілеві гриби, мікроскопічні водорості).
Людина використовував Б., ще не знаючи про їхнє існування. За допомогою заквасок, що містять Б., готували кисломолочні продукти, оцет, тісто і т.д. Вперше Б. побачив А. Левенгук - творець мікроскопа, досліджуючи рослинні настої і зубний наліт. До кінця 19 - початку 20 ст. було виділено велике число Б., що мешкають в грунті, воді, харчових продуктах і т.п., були відкриті багато видів хвороботворних Б. Класичні дослідження Л. Пастера в області фізіології Б. послужили основою для вивчення у них обміну речовин. Внесок у дослідження Б. внесли російські і радянські вчені С. Н. Виноградський , В. Л. Омелянський , Б. Л. Ісаченко , Які з'ясували роль Б. в круговороті речовин в природі, який робить можливим життя на Землі. Цей напрямок в мікробіології нерозривно пов'язане з розвитком геології, біогеохімії, ґрунтознавства, з вченням В. І. Вернадського про біосфері .
Морфологія і систематика Б. Розмір, форма, будова, рухливість Б. Діаметр кулястих Б. зазвичай дорівнює 1-2 мкм, ширина паличковидних форм коливається від 0,4 до 0,8 мкм, довжина дорівнює 2-5 мкм. Рідше зустрічаються дуже великі Б. Так, серобактерія Thiophysa macrophysa має 20 мкм в діаметрі, нитки інших серобактерий (Beggiatoa) видно неозброєним оком. Є також дуже дрібні Б., наприклад Bdellovibrio, що паразитують на Б. звичайних розмірів. Деякі Б., наприклад збудники плевропневмонії рогатої худоби, настільки дрібні, що невидимі в оптичний мікроскоп. Кулясті бактерії називаються кокками , Якщо ж вони розташовуються попарно, - диплококами (Рис. 1). Якщо коки розмножуються поперечним діленням і після ділення залишаються з'єднаними, утворюючи ланцюжки, то їх називають стрептококками (Рис. 2, 12). При діленні клітин в трьох взаємно перпендикулярних напрямках утворюються пакети кліток, типові для сарцин (Рис. 3). При розподілі коків в різних площинах виникають скупчення клітин у вигляді грона винограду, що характерно для стафілококів (Рис. 4). Паличкоподібні Б., що утворюють спори, називаються бацилами . Паличкоподібні форми можуть мати «обрубані» або опуклі кінці (рис. 5) і розташовуються окремо або, рідше, у вигляді ланцюжка (рис. 6). Б., що утворюють довгі нитки, - нитчасті бактерії , Мешкають переважно у воді. Б. у формі коми - вібріони (Рис. 7), покручені форми з грубими спіральними завитками - спірили (Рис. 8), з декількома рівномірними тонкими завитками - спірохети (Рис. 9).
Все Б. мають клітинну стінку (рис. 25, 26, 28). Вона виразно видно при приміщенні Б. у розчин кухонної солі; при цьому вміст клітини стискається і відстає від стінки - настає плазмоліз (Рис. 10). У ряду Б. стінка оточена слизової капсулою (рис. 27), присутність якої може бути встановлено при приміщенні таких Б. в розчин туші (рис. 11). При електронній мікроскопії видно, що клітинна стінка складається з декількох шарів (зазвичай трьох). До її складу входять мураміновая кислота, амінокислоти, ліпіди, глюкозамін і інші з'єднання. Хімічний склад клітинної стінки у різних систематичних груп, а також у Б., забарвлюються і не забарвлюються за Грамом, різний. Велику роль в обміні речовин грає цитоплазматична мембрана, яка перебуває під клітинною стінкою. У мембрані (рис. 25, 28) зосереджені численні ферментні системи бактеріальної клітини. У цитоплазмі є рибосоми , До складу яких входить РНК. Вміст нуклеїнових кислот у Б. коливається від 10 до 22% при різному ставленні РНК / ДНК (у кишкової палички воно дорівнює 2). За допомогою електронного мікроскопа встановлено присутність в клітці Б. ниток ДНК, що утворюють ядро, позбавлене оболонки, - т. Зв. нуклеоид (рис. 25, 26, 28). Будова ядра неоднаково у різних Б. Так, у «вищих», складніше організованих Б. (Myxobacteriales, Hyphomycrobiales), ядра легко можуть бути виявлені при мікроскопії забарвлених препаратів в оптичному мікроскопі (рис. 1). У багатьох Б. цитоплазма ущільнена на кінцях кліток і утворює забарвлюються полярні зерна (рис. 13; рис. 2). Клітини Б. містять запасні речовини: жирові включення (рис. 5), зерна глікогену, метахроматіна (рис. 4), гранульози (рис. 6), а також вакуолі, що містять рідину або газ. На відміну від грибів, Б. не містять мітохондрій , Що свідчить про більш примітивному будову Б. Багато Б. рухливі. Зазвичай такі форми мають довгі джгутики, що складаються з скорочувального білка (рис. 14). Завдяки хвилеподібним і спіральним рухам джгутиків клітка Б. переміщається. Види з одним джгутиком на полюсі клітини зв. монотріхі (рис. 15), клітини, що мають пучок джгутиків на кінці, - лофотрихи (рис. 16), Б., у яких джгутики розташовані по всій поверхні тіла, - перітріхі (рис. 17). У міксобактерій , Які також рухливі, джгутиків немає, і вони пересуваються в результаті набухання в довкіллі слизу, що виділяється клітинами (реактивний спосіб руху).
Життєвий цикл. Зміна морфології клітин Б. в часі дає уявлення про їх життєвому циклі. Так, багато аеробних і анаеробних Б. утворюють овальні або круглі блискучі спори . Такі види Б. називаються спороносними (або бацилами). Якщо суперечки великі і розташовуються в центрі клітини, то паличка набуває веретеноподібної форми (рис. 20); у інших видів спору розташовується на кінці палички, і тоді остання набуває форму булави (рис. 19) або барабанної палички. У багатьох спороносних бактерій діаметр спори невеликий, і тому при утворенні спори зберігається паличкоподібна форма Б. (рис. 18, 21). Надалі залишки вегетативної клітини руйнуються, і суперечки стає вільною (рис. 22). У кожній клітині утворюється тільки одна спору і, отже, спорообразование не можна розглядати як розмноження. Спори Б. дуже стійкі до дії високої температури і отруйних речовин. Потрапивши в сприятливе живильне середовище, спори проростають і з них виходять молоді паличкоподібні вегетативні клітини (рис. 23). Цикл розвитку Б. може бути різним. Так, мікобактерії розмножуються як діленням, так і брунькуванням (рис. 25). У міксобактерій вегетативні клітини стискаються, скорочуються і утворюють круглі або овальні мікроцисти (див. циста ), Які потім можуть проростати (рис. 24). Сполучені слизом мікроцисти утворюють тіла кулястої, грибовидной або Коралоподібних форми зеленого, рожевого або іншого кольору. В процесі зростання Б. можуть утворювати фільтрівні форми, що проходять через фільтри і дають в подальшому культури, подібні або тотожні з тими, в яких вони виникли.
Мінливість, фізіологічні і морфологічні властивості Б. можуть піддаватися змінам. Б. можуть втратити рухливість, здатність утворювати пігменти, давати суперечки, посилювати або зменшувати здатність до синтезу різних органічних сполук, змінювати форму і будову колоній на щільних поживних середовищах і т.д. Ці зміни можуть відбуватися мимовільно, т. Е. Безпрімененія відповідних зовнішніх впливів. Значно більше змінених форм виникає в результаті застосування мутагенів (ультрафіолетові промені, іонізуюча радіація, етиленімін або інших хімічних речовини). Кожна властивість Б. пов'язано з ДНК, т. Е. Контролюється відповідним геном . завдяки успіхам генетики мікроорганізмів встановлено місце розташування багатьох генів в нитки ДНК (див. Генетичні карти хромосом ). Виділивши ДНК з клітин-мутантів і додавши її в культуру іншого штаму, можна викликати (в результаті проникнення ДНК всередину клітин) спадкові зміни, називаються трансформацією . За допомогою мутагенів можуть бути отримані мутанти, цінні в практичному відношенні, в тому числі утворюють більшу кількість різних антибіотиків, амінокислот, вітамінів та інших біологічно активних речовин. За допомогою мутантів Б. були розшифровані шляху біосинтезу різних органічних сполук. Поступово змінюючи середовище проживання Б., можна адаптувати їх до нових умов існування. Так були отримані форми, стійкі до різних отрут, що розвиваються при незвичайній температурі або реакції середовища, і т.п .; так з'являються стійкі до деяких антибіотиків форми хвороботворних Б.
Систематика. Для з'ясування систематич. положення Б. визначають їх розміри, морфологію клітин, характер зростання чистої культури на різних питати. середовищах, форму, колір і характер поверхні колоній, що виростають на щільних середовищах. Встановлюють також характер розрідження Б. желатин (рис. 30), здатність їх згортати молоко, зброджувати різні вуглеводи, відновлювати нітрати, ооразовивать аміак, сірководень і індол при розкладанні білків і т.п. Надалі, маючи характеристику виділеної культури, визначають її систематичне положення. Б. підрозділяють на три класи. Перший клас - Eubacteria - об'єднує Б., що мають щільну клітинну стінку і не утворюють плодових тіл. В цьому класі розрізняють наступні порядки: 1) Eubacteriales - одноклітинні коки, неветвящиеся палички і спірально звиті форми; до цього порядку відносяться всі неспороносні і спороносні Б., фото-синтезують Б., спірохети та ін .; 2) Trichobacteriales - багатоклітинні нитчасті Б. з поперечними перегородками; 3) Ferribacteriales - одноклітинних нитчасті автотрофні железобактерии; 4) Thiobacteriales - одноклітинні автотрофні серобактерии. Другий клас - Myxobacteria - об'єднує Б. з тонкою клітинною стінкою і реактивним характером руху, що утворюють мікроцисти і плодові тіла різної форми. До третього класу - Hyphomicrobiales - відносять клітки, що дають довгі нитки, на кінцях яких утворюються нирки; відокремилися нирки рухливі. Сов. систематики виділяють на підставі еволюційних (філогенезі) даних великі групи, які об'єднують родинні форми. Так, наприклад, розгалужені мікобактерії не виділяються в самостійну групу, а об'єднані з актиноміцетами.
Фізіологія Б. Зростання, розмноження, розвиток. Після поділу Б. кожна з двох дочірніх бактеріальних клітин починає рости і досягає розмірів материнської. У цьому випадку говорять про зростання окремої клітини. Розмноження клітин, складових популяцію , Призводить до збільшення загального числа клітин. У цьому випадку говорять про зростання культури. При зростанні культури в рідкому поживному середовищі остання стає каламутною; чим більше клітин в культурі, тим вона мутніше. Про інтенсивність росту судять на підставі підрахунку клітин в 1 мл культури за допомогою мікроскопа або визначають за допомогою нефелометра ступінь каламутності живильного середовища. Визначаючи кількість клітин в різні періоди зростання культури, можна отримати криву зростання, що відображає кілька фаз: спочатку клітини не розмножуються, потім починають ділитися, причому швидкість розмноження весь час зростає; далі настає фаза, для якої характерна постійна швидкість ділення клітин; потім ця швидкість зменшується і настає відмирання клітин. Для отримання максимальної кількості клітин Б. вирощують в умовах т. Н. проточною культури; при цьому з посудини, в якому розмножуються Б., витікає певний обсяг культури; одночасно в судину додається в такій же кількості свіжа стерильна живильне середовище. При розмноженні Б. не в проточних, а в стаціонарних умовах відбувається зміна живильного середовища і накопичення в ній продуктів життєдіяльності Б., унаслідок чого змінюються і їх фізіологічні особливості. Так, молоді клітини Clostridium acetobutylicum не здатні утворювати ацетон; це властивість вони набувають в старішої культурі. Якщо спороносних Б. вирощувати в умови проточної культури, вони ділитимуться, але не даватимуть суперечку. При вирощуванні Б. на щільних поживних середовищах вони утворюють скупчення клітин різних розмірів, форми, кольору, звані колоніями.
Харчування. До складу клітин Б. входять ті ж біогенні елементи і мікроелементи, що і до складу клітин вищих рослин і тварин. Це С, N, О, Н, S, Р, К, Mg, Ca, Cl, Fe н ін. Крім білка, вуглеводів і жирів, Б. містять також РНК і велика кількість ДНК. Всі ці речовини можуть бути синтезовані тільки з речовин, що містяться в навколишньому середовищі. Як правило, через напівпроникну клітинну стінку і цитоплазматичну мембрану всередину Б. проходять тільки розчинні речовини. Під дією гідролітичних ферментів, що надходять з бактеріальних клітин назовні, відбувається розкладання складніших речовин (наприклад, крохмалю, целюлози) з утворенням розчинних продуктів (наприклад, моносахаров), засвоюваних Б. Як джерело азоту Б. можуть засвоювати білки, амінокислоти, амонійні солі , нітрати. Різні види Б. здатні утилізувати різні джерела азоту. Раніше вважали, що деякі патогенні (хвороботворні) і молочнокислі Б. можуть розвиватися лише в живильних середовищах з білками. Надалі з'ясувалося, що джерелом азоту для таких Б. можуть служити амонійні солі. Існує багато видів Б. з різних систематичних груп, які здатні засвоювати не тільки азот тих чи інших азотовмісних речовин, але і фіксувати азот атмосфери. До таких азотфиксирующим мікроорганізмам відносяться азотобактерии, мікобактерії, пурпурні фотосинтезирующие Б., а також бульбочкові бактерії (Див. Азотфіксация ). Джерелами мінерального живлення для Б. служать солі Р, S, Cl, К, Fe, Na, Ca; багато видів потребують також мікроелементів (Mo, Мn, Си, В, V і ін.). Для розмноження Б. необхідні також чинники зростання мікроорганізмів, до яких належать вітаміни групи В, біотин, пантотенова кислота, фолієва кислота та ін. Б., здатні синтезувати ці речовини, називаються ауксоавтотрофамі. До них відносяться псевдомоноси і багато інших неспороносні Б. При вирощуванні Б., не здатних до синтезу чинників зростання, їх додають в живильне середовище. Такі Б. називаються а у ксогетеротрофам і, до них відносяться, наприклад, різні молочнокислі Б. Джерелами вуглецю для Б. можуть служити вуглеводи, спирти, органічні кислоти, лігнін, хітин, вуглеводні, жири та ін. Здатність засвоювати вуглець з тих чи інших джерел у різних видів Б. варіює, і цим користуються для цілей систематики. Б., що засвоюють вуглець органічних сполук, називаються гетеротрофних. Б., що засвоюють вуглець вуглекислого газу атмосфери, називаються автотрофними. Види, що використовують для фіксації вуглекислоти енергію сонячних променів, наз. фотоавтотрофамі. Група Б., одержуючих енергію в результаті окислення таких неорганічних речовин, як аміак, нітрити, сірка, водень і ін., Здатних засвоювати вуглекислоту за рахунок енергії, що звільняється при окисленні вказаних неорганічних з'єднань, називаються хемоавтотрофов і, а сам процес асиміляції двоокису вуглецю, відкритий видатним російським мікробіологом С. Н. Виноградским, - хемосинтезом .
бактерії
Бакта е рії (грец. Bakterion - паличка), велика група (тип) мікроскопічних, переважно одноклітинних організмів, що володіють клітинною стінкою, що містять багато дезоксирибонуклеїнової кислоти (ДНК), що мають примітивне ядро, позбавлене видимих хромосом і оболонки, що не містять, як правило, хлорофілу і пластид, що розмножуються поперечним діленням (рідше перетяжкою або брунькуванням). Переважна кількість видів Б. має паличкоподібну форму. Однак до Б. відносять також мікроорганізми, що мають кулясту, ниткоподібну або звиту форму. Б. різноманітні по своїй фізіології, біохімічно дуже активні і поширені в грунті, воді, грунті водоймищ і пр. Б. не уявляють єдиної групи, а виникли різними шляхами. Деякі Б. (наприклад, нитчасті, азотобактер і ін.) Близькі до синезеленим водоростям , Ін. Б. споріднені променистим грибкам - актиноміцетам , Спірохети і деякі ін. Б. мають схожість з одноклітинними тваринами - найпростішими .
Б. беруть участь в круговороті речовин в природі, деякі з них викликають захворювання людини, тварин або рослин, застосовуються в різних галузях мікробіологічної промисловості. Науку, що вивчає Б., називають бактериологией . Це складова частина ширшої дисципліни - мікробіології , В завдання якої входить вивчення всіх сторін життєдіяльності не тільки Б., але і інших мікроорганізмів (дріжджі, цвілеві гриби, мікроскопічні водорості).
Людина використовував Б., ще не знаючи про їхнє існування. За допомогою заквасок, що містять Б., готували кисломолочні продукти, оцет, тісто і т.д. Вперше Б. побачив А. Левенгук - творець мікроскопа, досліджуючи рослинні настої і зубний наліт. До кінця 19 - початку 20 ст. було виділено велике число Б., що мешкають в грунті, воді, харчових продуктах і т.п., були відкриті багато видів хвороботворних Б. Класичні дослідження Л. Пастера в області фізіології Б. послужили основою для вивчення у них обміну речовин. Внесок у дослідження Б. внесли російські і радянські вчені С. Н. Виноградський , В. Л. Омелянський , Б. Л. Ісаченко , Які з'ясували роль Б. в круговороті речовин в природі, який робить можливим життя на Землі. Цей напрямок в мікробіології нерозривно пов'язане з розвитком геології, біогеохімії, ґрунтознавства, з вченням В. І. Вернадського про біосфері .
Морфологія і систематика Б. Розмір, форма, будова, рухливість Б. Діаметр кулястих Б. зазвичай дорівнює 1-2 мкм, ширина паличковидних форм коливається від 0,4 до 0,8 мкм, довжина дорівнює 2-5 мкм. Рідше зустрічаються дуже великі Б. Так, серобактерія Thiophysa macrophysa має 20 мкм в діаметрі, нитки інших серобактерий (Beggiatoa) видно неозброєним оком. Є також дуже дрібні Б., наприклад Bdellovibrio, що паразитують на Б. звичайних розмірів. Деякі Б., наприклад збудники плевропневмонії рогатої худоби, настільки дрібні, що невидимі в оптичний мікроскоп. Кулясті бактерії називаються кокками , Якщо ж вони розташовуються попарно, - диплококами (Рис. 1). Якщо коки розмножуються поперечним діленням і після ділення залишаються з'єднаними, утворюючи ланцюжки, то їх називають стрептококками (Рис. 2, 12). При діленні клітин в трьох взаємно перпендикулярних напрямках утворюються пакети кліток, типові для сарцин (Рис. 3). При розподілі коків в різних площинах виникають скупчення клітин у вигляді грона винограду, що характерно для стафілококів (Рис. 4). Паличкоподібні Б., що утворюють спори, називаються бацилами . Паличкоподібні форми можуть мати «обрубані» або опуклі кінці (рис. 5) і розташовуються окремо або, рідше, у вигляді ланцюжка (рис. 6). Б., що утворюють довгі нитки, - нитчасті бактерії , Мешкають переважно у воді. Б. у формі коми - вібріони (Рис. 7), покручені форми з грубими спіральними завитками - спірили (Рис. 8), з декількома рівномірними тонкими завитками - спірохети (Рис. 9).
Все Б. мають клітинну стінку (рис. 25, 26, 28). Вона виразно видно при приміщенні Б. у розчин кухонної солі; при цьому вміст клітини стискається і відстає від стінки - настає плазмоліз (Рис. 10). У ряду Б. стінка оточена слизової капсулою (рис. 27), присутність якої може бути встановлено при приміщенні таких Б. в розчин туші (рис. 11). При електронній мікроскопії видно, що клітинна стінка складається з декількох шарів (зазвичай трьох). До її складу входять мураміновая кислота, амінокислоти, ліпіди, глюкозамін і інші з'єднання. Хімічний склад клітинної стінки у різних систематичних груп, а також у Б., забарвлюються і не забарвлюються за Грамом, різний. Велику роль в обміні речовин грає цитоплазматична мембрана, яка перебуває під клітинною стінкою. У мембрані (рис. 25, 28) зосереджені численні ферментні системи бактеріальної клітини. У цитоплазмі є рибосоми , До складу яких входить РНК. Вміст нуклеїнових кислот у Б. коливається від 10 до 22% при різному ставленні РНК / ДНК (у кишкової палички воно дорівнює 2). За допомогою електронного мікроскопа встановлено присутність в клітці Б. ниток ДНК, що утворюють ядро, позбавлене оболонки, - т. Зв. нуклеоид (рис. 25, 26, 28). Будова ядра неоднаково у різних Б. Так, у «вищих», складніше організованих Б. (Myxobacteriales, Hyphomycrobiales), ядра легко можуть бути виявлені при мікроскопії забарвлених препаратів в оптичному мікроскопі (рис. 1). У багатьох Б. цитоплазма ущільнена на кінцях кліток і утворює забарвлюються полярні зерна (рис. 13; рис. 2). Клітини Б. містять запасні речовини: жирові включення (рис. 5), зерна глікогену, метахроматіна (рис. 4), гранульози (рис. 6), а також вакуолі, що містять рідину або газ. На відміну від грибів, Б. не містять мітохондрій , Що свідчить про більш примітивному будову Б. Багато Б. рухливі. Зазвичай такі форми мають довгі джгутики, що складаються з скорочувального білка (рис. 14). Завдяки хвилеподібним і спіральним рухам джгутиків клітка Б. переміщається. Види з одним джгутиком на полюсі клітини зв. монотріхі (рис. 15), клітини, що мають пучок джгутиків на кінці, - лофотрихи (рис. 16), Б., у яких джгутики розташовані по всій поверхні тіла, - перітріхі (рис. 17). У міксобактерій , Які також рухливі, джгутиків немає, і вони пересуваються в результаті набухання в довкіллі слизу, що виділяється клітинами (реактивний спосіб руху).
Життєвий цикл. Зміна морфології клітин Б. в часі дає уявлення про їх життєвому циклі. Так, багато аеробних і анаеробних Б. утворюють овальні або круглі блискучі спори . Такі види Б. називаються спороносними (або бацилами). Якщо суперечки великі і розташовуються в центрі клітини, то паличка набуває веретеноподібної форми (рис. 20); у інших видів спору розташовується на кінці палички, і тоді остання набуває форму булави (рис. 19) або барабанної палички. У багатьох спороносних бактерій діаметр спори невеликий, і тому при утворенні спори зберігається паличкоподібна форма Б. (рис. 18, 21). Надалі залишки вегетативної клітини руйнуються, і суперечки стає вільною (рис. 22). У кожній клітині утворюється тільки одна спору і, отже, спорообразование не можна розглядати як розмноження. Спори Б. дуже стійкі до дії високої температури і отруйних речовин. Потрапивши в сприятливе живильне середовище, спори проростають і з них виходять молоді паличкоподібні вегетативні клітини (рис. 23). Цикл розвитку Б. може бути різним. Так, мікобактерії розмножуються як діленням, так і брунькуванням (рис. 25). У міксобактерій вегетативні клітини стискаються, скорочуються і утворюють круглі або овальні мікроцисти (див. циста ), Які потім можуть проростати (рис. 24). Сполучені слизом мікроцисти утворюють тіла кулястої, грибовидной або Коралоподібних форми зеленого, рожевого або іншого кольору. В процесі зростання Б. можуть утворювати фільтрівні форми, що проходять через фільтри і дають в подальшому культури, подібні або тотожні з тими, в яких вони виникли.
Мінливість, фізіологічні і морфологічні властивості Б. можуть піддаватися змінам. Б. можуть втратити рухливість, здатність утворювати пігменти, давати суперечки, посилювати або зменшувати здатність до синтезу різних органічних сполук, змінювати форму і будову колоній на щільних поживних середовищах і т.д. Ці зміни можуть відбуватися мимовільно, т. Е. Безпрімененія відповідних зовнішніх впливів. Значно більше змінених форм виникає в результаті застосування мутагенів (ультрафіолетові промені, іонізуюча радіація, етиленімін або інших хімічних речовини). Кожна властивість Б. пов'язано з ДНК, т. Е. Контролюється відповідним геном . завдяки успіхам генетики мікроорганізмів встановлено місце розташування багатьох генів в нитки ДНК (див. Генетичні карти хромосом ). Виділивши ДНК з клітин-мутантів і додавши її в культуру іншого штаму, можна викликати (в результаті проникнення ДНК всередину клітин) спадкові зміни, називаються трансформацією . За допомогою мутагенів можуть бути отримані мутанти, цінні в практичному відношенні, в тому числі утворюють більшу кількість різних антибіотиків, амінокислот, вітамінів та інших біологічно активних речовин. За допомогою мутантів Б. були розшифровані шляху біосинтезу різних органічних сполук. Поступово змінюючи середовище проживання Б., можна адаптувати їх до нових умов існування. Так були отримані форми, стійкі до різних отрут, що розвиваються при незвичайній температурі або реакції середовища, і т.п .; так з'являються стійкі до деяких антибіотиків форми хвороботворних Б.
Систематика. Для з'ясування систематич. положення Б. визначають їх розміри, морфологію клітин, характер зростання чистої культури на різних питати. середовищах, форму, колір і характер поверхні колоній, що виростають на щільних середовищах. Встановлюють також характер розрідження Б. желатин (рис. 30), здатність їх згортати молоко, зброджувати різні вуглеводи, відновлювати нітрати, ооразовивать аміак, сірководень і індол при розкладанні білків і т.п. Надалі, маючи характеристику виділеної культури, визначають її систематичне положення. Б. підрозділяють на три класи. Перший клас - Eubacteria - об'єднує Б., що мають щільну клітинну стінку і не утворюють плодових тіл. В цьому класі розрізняють наступні порядки: 1) Eubacteriales - одноклітинні коки, неветвящиеся палички і спірально звиті форми; до цього порядку відносяться всі неспороносні і спороносні Б., фото-синтезують Б., спірохети та ін .; 2) Trichobacteriales - багатоклітинні нитчасті Б. з поперечними перегородками; 3) Ferribacteriales - одноклітинних нитчасті автотрофні железобактерии; 4) Thiobacteriales - одноклітинні автотрофні серобактерии. Другий клас - Myxobacteria - об'єднує Б. з тонкою клітинною стінкою і реактивним характером руху, що утворюють мікроцисти і плодові тіла різної форми. До третього класу - Hyphomicrobiales - відносять клітки, що дають довгі нитки, на кінцях яких утворюються нирки; відокремилися нирки рухливі. Сов. систематики виділяють на підставі еволюційних (філогенезі) даних великі групи, які об'єднують родинні форми. Так, наприклад, розгалужені мікобактерії не виділяються в самостійну групу, а об'єднані з актиноміцетами.
Фізіологія Б. Зростання, розмноження, розвиток. Після поділу Б. кожна з двох дочірніх бактеріальних клітин починає рости і досягає розмірів материнської. У цьому випадку говорять про зростання окремої клітини. Розмноження клітин, складових популяцію , Призводить до збільшення загального числа клітин. У цьому випадку говорять про зростання культури. При зростанні культури в рідкому поживному середовищі остання стає каламутною; чим більше клітин в культурі, тим вона мутніше. Про інтенсивність росту судять на підставі підрахунку клітин в 1 мл культури за допомогою мікроскопа або визначають за допомогою нефелометра ступінь каламутності живильного середовища. Визначаючи кількість клітин в різні періоди зростання культури, можна отримати криву зростання, що відображає кілька фаз: спочатку клітини не розмножуються, потім починають ділитися, причому швидкість розмноження весь час зростає; далі настає фаза, для якої характерна постійна швидкість ділення клітин; потім ця швидкість зменшується і настає відмирання клітин. Для отримання максимальної кількості клітин Б. вирощують в умовах т. Н. проточною культури; при цьому з посудини, в якому розмножуються Б., витікає певний обсяг культури; одночасно в судину додається в такій же кількості свіжа стерильна живильне середовище. При розмноженні Б. не в проточних, а в стаціонарних умовах відбувається зміна живильного середовища і накопичення в ній продуктів життєдіяльності Б., унаслідок чого змінюються і їх фізіологічні особливості. Так, молоді клітини Clostridium acetobutylicum не здатні утворювати ацетон; це властивість вони набувають в старішої культурі. Якщо спороносних Б. вирощувати в умови проточної культури, вони ділитимуться, але не даватимуть суперечку. При вирощуванні Б. на щільних поживних середовищах вони утворюють скупчення клітин різних розмірів, форми, кольору, звані колоніями.
Харчування. До складу клітин Б. входять ті ж біогенні елементи і мікроелементи, що і до складу клітин вищих рослин і тварин. Це С, N, О, Н, S, Р, К, Mg, Ca, Cl, Fe н ін. Крім білка, вуглеводів і жирів, Б. містять також РНК і велика кількість ДНК. Всі ці речовини можуть бути синтезовані тільки з речовин, що містяться в навколишньому середовищі. Як правило, через напівпроникну клітинну стінку і цитоплазматичну мембрану всередину Б. проходять тільки розчинні речовини. Під дією гідролітичних ферментів, що надходять з бактеріальних клітин назовні, відбувається розкладання складніших речовин (наприклад, крохмалю, целюлози) з утворенням розчинних продуктів (наприклад, моносахаров), засвоюваних Б. Як джерело азоту Б. можуть засвоювати білки, амінокислоти, амонійні солі , нітрати. Різні види Б. здатні утилізувати різні джерела азоту. Раніше вважали, що деякі патогенні (хвороботворні) і молочнокислі Б. можуть розвиватися лише в живильних середовищах з білками. Надалі з'ясувалося, що джерелом азоту для таких Б. можуть служити амонійні солі. Існує багато видів Б. з різних систематичних груп, які здатні засвоювати не тільки азот тих чи інших азотовмісних речовин, але і фіксувати азот атмосфери. До таких азотфиксирующим мікроорганізмам відносяться азотобактерии, мікобактерії, пурпурні фотосинтезирующие Б., а також бульбочкові бактерії (Див. Азотфіксация ). Джерелами мінерального живлення для Б. служать солі Р, S, Cl, К, Fe, Na, Ca; багато видів потребують також мікроелементів (Mo, Мn, Си, В, V і ін.). Для розмноження Б. необхідні також чинники зростання мікроорганізмів, до яких належать вітаміни групи В, біотин, пантотенова кислота, фолієва кислота та ін. Б., здатні синтезувати ці речовини, називаються ауксоавтотрофамі. До них відносяться псевдомоноси і багато інших неспороносні Б. При вирощуванні Б., не здатних до синтезу чинників зростання, їх додають в живильне середовище. Такі Б. називаються а у ксогетеротрофам і, до них відносяться, наприклад, різні молочнокислі Б. Джерелами вуглецю для Б. можуть служити вуглеводи, спирти, органічні кислоти, лігнін, хітин, вуглеводні, жири та ін. Здатність засвоювати вуглець з тих чи інших джерел у різних видів Б. варіює, і цим користуються для цілей систематики. Б., що засвоюють вуглець органічних сполук, називаються гетеротрофних. Б., що засвоюють вуглець вуглекислого газу атмосфери, називаються автотрофними. Види, що використовують для фіксації вуглекислоти енергію сонячних променів, наз. фотоавтотрофамі. Група Б., одержуючих енергію в результаті окислення таких неорганічних речовин, як аміак, нітрити, сірка, водень і ін., Здатних засвоювати вуглекислоту за рахунок енергії, що звільняється при окисленні вказаних неорганічних з'єднань, називаються хемоавтотрофов і, а сам процес асиміляції двоокису вуглецю, відкритий видатним російським мікробіологом С. Н. Виноградским, - хемосинтезом .
Дихання и обмін Речовини. Синтез Речовини, что входять до складу бактеріальної Клітини, ее рухлівість и інші процеси супроводжуються витрати ЕНЕРГІЇ. Більшість Б. отрімує Енергію Шляхом окислення органічніх Речовини, хемоавтотрофні - в результате окислення неорганічніх Сполука, фотосинтезирующие Б. Використовують Енергію Сонячних променів. Б., здатні рости только в прісутності кисня, назіваються аеробамі , Що ростуть во время відсутності кисня, - анаероби . При аеробних діханні відбувається окислення органічніх Сполука з віділенням вуглекислий газу. Если ж окислення идет не до кінця, то в середовіщі накопічуються проміжні продукти. Такі процеси назіваються окіслювальнімі бродіння (например, уксуснокислое бродіння). Розкладання органічніх Речовини в анаеробних условиях Із звільненням ЕНЕРГІЇ назіваються бродінням . При зброджуванні вуглеводів різнімі Б. могут утворюватіся: молочна або масляна кислота, етиловий, пропіловій або бутиловий спирт, ацетон и інші Речовини. Ряд біохімічніх процесів (гліколіз, транспорт електронів, цикл Кребса, синтез амінокіслот, білків, нуклеїнових кислот и ін.) Протікає у Б. почти так само, як в клітінах рослин и тварин. Специфічне особливості обміну речовин Б. - висока біохімічна активність, здатність окислювати неорганічні сполуки сірки, азоту (аміак) і ін., Синтезувати білок, використовуючи в якості вихідного продукту фенол, метан та інші вуглеводні, окисляти водень, фіксувати азот атмосфери, синтезувати ферменти, розкладають целюлозу або лігнін, утворювати метан з вуглекислоти і водню та ін. - виключно цінні в практичному відношенні.
Екологія Б. Поширення. Б. відносяться до космополітів. Одні і ті ж види Б. можна знайти на всіх материках, т. Е. Майже повсюдно. Кількість їх в грунті, воді та інших середовищах визначають прямим підрахунком клітин в пофарбованому препараті або посівом на різні поживні середовища. В 1 г грунту містяться сотні тисяч або мільйони бактерій; в 1 мл води - десятки або сотні клітин. Сильний вплив на бактеріальну мікрофлору надають екологічні умови. Так, окультурені грунту не тільки містять більше Б., чим, наприклад, грунти пустель, але і розрізняються за видовим складом мікрофлори. Сучасній мікробіології відомо не більше 1/10 частини Б., що існують в природі. Застосування методів капілярної і електронної мікроскопії препаратів грунту дозволило виявити багато нових видів Б. Розвиваючись в самих різних екологічних умовах, Б. в процесі еволюції пристосувалися до них. Так виникли термофільні Б., що мешкають у воді гарячих джерел, в розігрівають купах торфу або гною, психрофільні форми, що живуть при низькій температурі в воді полярних морів, галофільні Б., здатні розмножуватися в середовищі, що містить до 20% кухонної солі, ацидофільні та алкалофільних Б., зростаючі в дуже кислою або сильно лужному середовищі, і т.д. Широке поширення в природі певних джерел вуглецю або азоту привело в ході еволюції до фізіологічної конвергенції , Т. Е. Появі у представників різних систематичних груп Б. здатності засвоювати біогенний елемент з одного джерела. Так, Б., здатні фіксувати атмосферний азот, належать до різних класів, порядків і сімейств; А; здатністю утилізувати целюлозу мають багато Б., далекі в систематичному відношенні. Між різними видами. Б., з одного боку, і іншими мікроорганізмами, рослинами або тваринами - з іншого, можуть існувати як антагоністичні (див. антагонізм ), Так і симбіотичні (див. симбіоз ) Відносини. Деякі Б. утворюють пігменти, антибіотики або органічні кислоти, що пригнічують життєдіяльність інших Б., грибів, водоростей, одноклітинних і деяких клітин багатоклітинних тварин. Бактеріальні віруси - бактеріофаги - проникають всередину Б. і, розмножуючись в них, викликають загибель і лізис мікроорганізмів. При симбіотичних, т. Е. Заснованих на взаємній користі, стосунках один вид Б. може споживати продукти життєдіяльності іншого виду, накопичення яких в культуральній рідині гальмує зростання останнього. У свою чергу симбіонт може виділяти в середу додаткові чинники зростання, необхідні іншого виду. Б., що мешкають в кишечнику тварин або людини і харчуються за рахунок вмісту кишечника, утворюють ферменти, необхідні для травлення, а також ряд речовин, вкрай важливих для життя господаря (незамінні амінокислоти, різні вітаміни та ін.).
Роль Б. в круговороті речовин в природі. Мінералізуя рослинні і тваринні залишки, мікроорганізми беруть участь у кругообігу всіх хімічних елементів, що входять до складу живих клітин. Так, джерелом вуглецю для вищих рослин і хемоавтотрофні Б. служить вуглекислота атмосфери, що фіксується в процесі фото- або хемосинтезу. біомаса рослин і тварин розкладається мікроорганізмами, здатними утилізувати целюлозу, пентози, крохмаль, лігнін, пектинові речовини, в кінцевому підсумку до вуглекислоти і води (рис. 31).
Так само велика роль мікроорганізмів, в тому числі Б., в кругообігу азоту. Тварини, харчуючись рослинами, синтезують білок та інші азотовмісні продукти свого тіла за рахунок білка рослин. При мінералізації тваринного і рослинного білка гнильні Б. утворюють аміак, який окислюється нітріфіцірующімі Б. в нітрити і потім в нітрати. Як амонійні солі, так і нітрати служать джерелом азотистого живлення для вищих рослин, що синтезують при цьому білки свого тіла (рис. 32). Мінераліз здатність Б. забезпечує круговорот і інших біогенних елементів. Руйнуючи органічні сполуки фосфору (нуклеїнові кислоти і ін.), Вони збагачують мінеральними з'єднаннями фосфору водоймища і грунт. Під впливом Б. відбувається мінералізація і органічних сполук сірки. Сірчані Б. можуть окислювати сірководень, сірку або деякі її сполуки до сірчаної кислоти, інші здатні відновлювати сульфати з утворенням сірководню. Б. здійснюють окислення заліза і марганцю, відкладення солей кальцію, окислення метану і водню, руйнування гірських порід продуктами життєдіяльності і ін. Все це дозволяє вважати Б. потужними геологічними діячами.
Практичне значення. Б. служать улюбленими об'єктами для вирішення загальних питань генетики, біохімії, біофізики, космічної біології і ін. Культури Б. застосовуються для кількісного визначення амінокислот, вітамінів, антибіотиків. Родючість ґрунтів значною мірою пов'язано з життєдіяльністю Б., минерализующих рослинні і тваринні залишки з утворенням з'єднань, що засвоюються с.-г. рослинами. Разом з тим, синтезуючи живу речовину клітин, Б. накопичують великі кількості органічних сполук в грунті. У верхніх шарах окультуреній грунту на площі в 1 га міститься декілька т бактеріальних клітин. Живучи в грунті азотфіксуючі Б. збагачують грунт азотом. Виключно велика роль бульбочкових бактерій, які фіксують газоподібний азот. Зараження насіння бобових рослин нитрагином - препаратом, що містить клітини бульбочкових Б. (див. бактеріальні добрива ), Підвищує урожай рослин і накопичення азоту в грунті. За допомогою Б., що зброджують пектинові речовини, здійснюють мочку льону, конопель, кенафа та інших луб'яних культур. Різні види Б. застосовують при отриманні з молока кисломолочних продуктів, масла і сиру.
У мікробіологічної промисловості за допомогою соответстветствующих видів Б. отримують з крахмалсодержащего або іншої сировини молочну кислоту, ацетон, етиловий, бутиловий та інші спирти, кровозамінник декстран, діацетил, антибіотики (граміцидин і ін.), Вітаміни, амінокислоти та ін. Особливо широко застосовуються Б . для отримання ферментних препаратів (амілази, протеази і ін.). В результаті розмноження Б., створюючих молочну кислоту з вуглеводів, квашена капуста, силос, солоні огірки не гниють, тому що кисла реакція заважає розвитку гнильних Б. Окисляющие сірку Б. застосовують для бактеріального вилуговування міді і ряду інших металів з містять їх порід. Помістивши здатні засвоювати газоооразние вуглеводні Б. у судини, що зариваються потім в грунт, можна на підставі зростання Б. укладати, є чи в цій місцевості нафта або природний газ. З багатьма видами Б. доводиться вести серйозну боротьбу, оберігаючи від псування і руйнування ними зерно, овочі, фрукти, всі харчові продукти, різні види сировини, матеріалів і виробів (текстиль, картон, канати, риболовні сіті, ізоляція кабелю і мн. Ін. ). Багато хвороб людини викликаються патогенними Б. До таких хвороб відносяться різні епідемічного захворювання (холера, черевний тиф, паратифи, чума, дифтерія, туляремія, бруцельоз), а також туберкульоз, зараження крові (сепсис), проказа, сифіліс та ін. У тварин Б . викликають сап, сибірську виразку, туберкульоз і ін. Багато хвороб як культурних, так і диких рослин викликають т. н. фітопатогенні Б. (див. Бактеріальні хвороби рослин ). Боротьба з хвороботворними Б. грунтується на асептики и антисептику , На застосуванні бактериостатических речовин и бактерицидних речовин (Див. антибіотики , хіміотерапія ).
Літ .: Красильников Н. А., Визначник бактерій і актиноміцетів, М.-Л., 1949; Ісаченко Б. Л., Вибрані праці, т. 1-2, М.-Л., 1951; Виноградський С. Н., Мікробіологія грунту, М., 1952; Кузнєцов С. І., Роль мікроорганізмів у кругообігу речовин в озерах, М., 1952; Імшенецький А. А., Мікробіологія целюлози, М., 1953; Омелянський В. Л., Вибрані праці, т. 1-2, М., 1953; Анатомія бактерій, пров. з англ., М., 1960; Работнова І. Л., Загальна мікробіологія, М., 1966; Clifton С. Є., Introduction to bacterial physiology, NY, 1957; Gunsalus 1. С. and Stanier RJ, The Bacteria, v. 1-5, NY, 1960; Stanier RJ, Dondoroff M., Adelberg Е. A., Themic-robial world, 2 ed., NY, 1963; Lamanna C., Mallette М. F., Basic bacteriology, 3 ed., Baltimore, 1965.
А. А. Імшенецький.
Рис. 27 до ст. Бактерії. Слизова капсула (а) у цисти азотобактера.
Рис. 23 до ст. Бактерії.
Включення запасного речовини - гранульози (забарвлюється йодом в синій колір) в клітинах анаеробної бактерії під час утворення нею суперечка.
Рис. 9 до ст. Бактерії. Спірохети.
Рис. 4 до ст. Бактерії. Стафілококи.
Рис. 30 до ст. Бактерії.
Рис. 25 до ст. Бактерії. Клітка золотистого стафілокока: а - клітинна стінка; б - цитоплазматична мембрана; в - нуклеоїд; г - мембранні структури всередині нуклеоида.
Рис. 2 до ст. Бактерії. Стрептококи.
Рис. 11 до ст. Бактерії.
Рис. 18 до ст. Бактерії.
Рис. 28 до ст. Бактерії. Клітка молочнокислої бактерії; а - внутрицитоплазматические мембранні структури (мезосоми); б - клітинна стінка; в - цитоплазматична мембрана.
Гігантські форми бактерій, що виникають при несприятливих умовах життя.
Рис. 19 до ст. Бактерії.
Клітини оцтовокислих бактерій із зернами метахроматіна.
Рис. 3 до ст. Бактерії. Сарціни.
Рис. 8 до ст. Бактерії. Спірілли.
Полярні зерна ( «шапочки»); чітко фарбуються у багатьох спороносних бактерій.
Рис. 15 до ст. Бактерії.
Рис. 17 до ст. Бактерії.
Рис. 13 до ст. Бактерії.
Клітини спороносной бактерії з великими краплями жиру.
Рис. 6 до ст. Бактерії. Бацили.
Рис. 20 до ст. Бактерії.
Рис. 29 до ст. Бактерії. Джгутик кишкової палички.
Рис. 26 до ст. Бактерії. Освіта поперечної перегородки в делящейся клітці гемолітичного стрептокока: а - утворюється перегородка; б - нуклеоїд; в - клітинна стінка.
Рис. 22 до ст. Бактерії.
Рис. 10 до ст. Бактерії. Плазмоліз.
Ядра в клітинах миксобактерии Polyangium; клітини розмножуються перетяжкою.
Рис. 1 до ст. Бактерії. Диплококи.
Рис. 32 до ст. Бактерії.
Рис. 12 до ст. Бактерії. Стрептококи.
Метахроматін в клітинах збудника дифтерії.
Рис. 5 до ст. Бактерії. Бацили.
Рис. 21 до ст. Бактерії.
Великі вигнуті клітини (спірили) із зернами метахроматіна.
Рис. 31 до ст. Бактерії.
Рис. 7 до ст. Бактерії. Вібріони.
Рис. 16 до ст. Бактерії.
Кислотостійкі палички прокази.
Рис. 24 до ст. Бактерії.
Рис. 14 до ст. Бактерії.

